Печінка, крупна залоза тваринного організму, що бере участь в процесах травлення, обміну речовин, кровообіги і що здійснює специфічні захисні і знешкоджуючі, ферментативні і видільні функції, направлені на підтримку постійності внутрішнього середовища організму.
Порівняльна морфологія. У безхребетних П. — травна залоза, що є вирощуванням середньої кишки, бере участь в переварюванні і всмоктуванні їжі; у ній відкладаються запасні живильні речовини, жири і вуглеводи. У багатьох безхребетних П. часто називається печінково-підшлунковою залозою (hepato-pancreas). У більшості молюсків П. масивна, часточкова, зазвичай парна; відкривається в шлунок 1—2 або багатьма протоками; клітки П. молюсків здатні до фагоцитозу . Серед членистоногих П. мають ракоподібні, мечехвости і більшість павукоподібних. П. ракоподібних — мішковидні вирости переднього відділу середньої кишки; виробляє фермент, що розщеплює клітковину. В павукоподібних П. представлена парними виростамі черевного відділу середньої кишки. Серед голкошкірих великі печінкові вирости шлунку є лише в морських лілій і зірок.
У хордових тварин і у людини П.— орган, в якому виробляється секрет, що бере участь в травленні (жовч), і здійснюються найважливіші процеси, пов'язані з обміном речовин в організмі. П. покривників — зазвичай залоза, що деревовидно гілкується; утворюється як вирощування шлунку, тісно прилягає до його стінки і відкривається в нього однією протокою. У ланцетника П. представлена мішковидним печінковим вирощуванням кишечника.
В хребетних П. закладається як черевне вирощування середньої кишки, яке, деревовидно розгалужуючись, набуває будови трубчастої залози. Просвіти кінцевих трубочок П. утворюють жовчні капіляри, по яких жовч стікає в крупніші печінкові протоки; окремі печінкові протоки зазвичай об'єднуються в загальний жовчна протока, що відкривається в дванадцятипалу кишку . Зазвичай з частини жовчної протоки утворюється жовчний міхур . Трубчаста будова П. зберігається протягом всього життя лише в деяких круглоротих (міксин). У міног, а також у риб і земноводних трубчаста будова П. частково порушується, оскільки між трубочками П. виникає поперечна щаблина — анастомози і між ними вростає сполучна тканина з кровоносними судинами і нервами. У плазунів, птиць і ссавців рясні анастомози перетворять трубчасту залозу в сітчасту. У міног і в деяких риб П.— нерозчленований орган, проте у більшості тварин в ній є права і ліва лопаті (жовчний міхур завжди пов'язаний з правою лопаттю). У деяких тварин (особливо серед ссавців) обидві лопаті можуть бути часточковими. У хижаків П. відносно більше, ніж в травоїдних. У риб і земноводних П.—крупнее, чим у плазунів, птиць і ссавців. Форма П. залежить від форми тіла тварини. В деяких земноводних, риб і ссавців П. тісно пов'язана з підшлунковою залозою, протоки якої відкриваються в жовчну протоку.
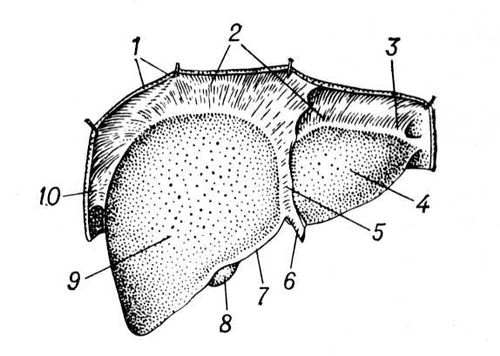
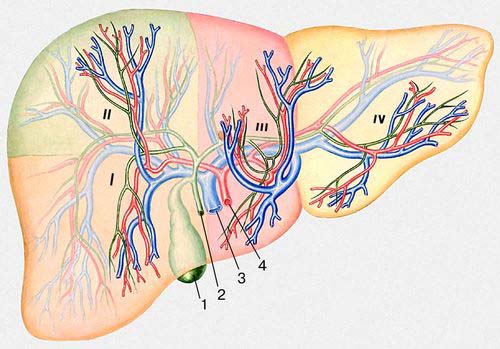
У людини П.— найкрупніша травна залоза. Розвивається на 3-му тижні внутріутробного життя плоду з вирощування епітелію слизистої оболонки дванадцятипалої кишки. У дорослої людини П. важить 1,5—2 кг (біля 1 / 50 маси тіла), має щільну консистенцію, але міняє форму і розміри як від дії з боку навколишніх органів, так і від функціональних навантажень. Зверху до П. прілежіт діафрагма (див. Грудобрюшная перешкода ), над якою розташовано серце і легені; знизу — шлунок, дванадцятипала кишка, частина поперечної ободової кишки, права брунька з наднирковою; ззаду — стравохід і хребетний стовп; спереду — передня стінка черевної порожнини. На П. розрізняють передневерхнюю, або діафрагмальну, і вісцелярну (ніжнюю) поверхні і краї: передній — гострий, задній — тупий. Передневерхняя опукла поверхня П. ділиться серповидною в'язкою на 2 долі — праву, велику, і ліву, меншу ( мал. 1 ). Нижня поверхня П. декілька увігнута. На ній розрізняють праву і ліву подовжні борозни і поперечну (називають воротами П.), які ділять П. на 4 долі: власне праву, ліву, хвостату, квадратну ( мал. 2 ). У правій подовжній борозні спереду лежить жовчний міхур, ззаду — ніжняя порожниста вена ; у лівій — спереду кругла в'язка П. (заросла пупкова вена), ззаду — венозна в'язка (заросле з'єднання пупкової вени з нижньою порожнистою веною). У ворота П. входять комірна вена, печінкова артерія, нерви; виходять — лімфатичні судини і печінкова протока, яка, з'єднуючись з протокою жовчного міхура, утворює желчевиносящий протока, що впадає в дванадцятипалу кишку. П. покрита серозною оболонкою (очеревиною ), за винятком ділянки, де вона прілежіт до діафрагмою зростається з нею. Серозна оболонка, переходячи с П. на сусідні органи, утворює в'язки: серповидну, праву і ліву вінцеві, сполучаючі П. з діафрагмою, а також що йдуть від воріт П. печінково-шлункову і ін. В'язки грають роль в утриманні П. на місці. Проте більше значення у фіксації П. мають: внутрішньочеревний тиск, обумовлююче більш менш щільне прилягання органів черевної порожнини один до одного і їх взаємну опору; нижня порожниста вена, щільно вростаюча своїми припливами (печінковими венами) в речовину П.; сила зчеплення між серозними оболонками П. і діафрагми в місцях їх зіткнення; сполучна тканина, що пов'язує П. з діафрагмою в місцях, не покритих очеревиною. П. розташована основною масою в правом підребер'я і через надчеревну область тягнеться в ліве підребер'я. Нижній кордон П. в нормі справа не виходить у дорослої людини з-під краю правої ребрової дуги. В новонароджених П. займає весь верхній відділ черевної порожнини і лівою долею стосується селезінки; нижній край П. незрідка доходить до пупка, у немовляти він виступає на 2—3 см з-під ребрового краю і лише до 4 років ховається за ним.
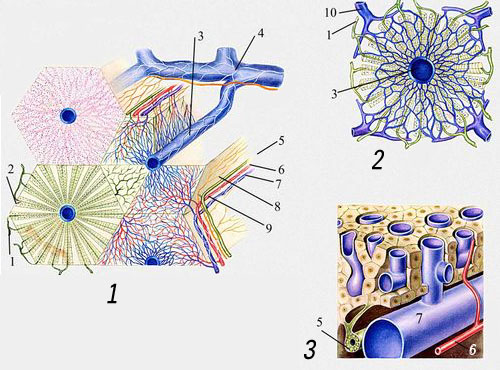
П. — складна трубчаста залоза. Під її серозною оболонкою розташовується соєдінітельнотканная (гліссонова) капсула, що містить еластичні волокна; у воротях П. капсула товщає і разом з кровоносними судинами проникає всередину П., розділяючи її на призматичні печінкові часточки розміром 0,5—2,0 мм (див. ілл. ). В середині кожної часточки проходіт центральна вена, від якої по радіусах у вигляді щаблини (балок), називається печінковими пластинками, розташовуються печінкові клітки. Печінкові пластинки в сукупності складають залізисту паренхіму П. Дольки П. складаються з тонких, але широких пластинок, анастомозуючих між собою і печінкових кліток, що складаються з одного шару. Між ними розташовуються жовчні капіляри, які, зливаючись, утворюють внутрі- і міжчасточкові жовчні протоки, складові печінкова протока.
Кровопостачання П. здійснюється по печінковій артерії, що приносить артеріальну кров, багату киснем, і комірній вені (див. Комірні системи ). Кров, що поступає в П. по комірній вені від шлунку, селезінки, кишечника, підшлункової залози і ін. органів черевної порожнини, містить деякі продукти переварювання білків, вуглеводів і частково жирів, різні хімічні речовини, що забезпечують фізіологічні функції П. Конечниє гілки печінкової артерії і комірної вени усередині часточок переходять в синусоїди, швидкість кровотоку в яких порівняно низька. Тут відбувається обмін між кров'ю і клітками П., після чого кров поступає в центральні вени, які з'єднуються і у вигляді 3—4 печінкових вен впадають в нижню порожнисту вену. Розгалужена капілярна мережа, поверхня якої досягає 400 м-коду ; забезпечує проходження через П. близько 2 тис. л крові в добу, причому 80% її поступає за системою комірної вени, а 20% — через печінкову артерію. Внутрішньочасточкові капіляри (синусоїди) в П., на відміну від звичайних капілярів, забезпечені, окрім ендотелію, ретикулярними клітками зірчастої форми (див. Купфера клітки ), які відносяться до ретікуло-ендотеліальній системі і володіють вираженою здібністю до фагоцитозу. Іннервіруєтся П. блукаючими нервами і гілками сонячного сплетення.
Фізіологія і біохімія. Фізіологічне значення П. визначається тим, що всмоктувані з кишечника в кров речовини проходять через П. і піддаються в ній хімічним змінам. П. бере участь в підтримці динамічної рівноваги багатьох речовин плазми крові (цукру, холестерину, білків крові, заліза, ретинолу, або вітаміну А, води). Через П. протікає в 1 мін близько 1,5 л крові, в їй звільняється 1 / 7 частина всієї енергії організму. Температура крові, що відтікала від неї, під час травлення збільшується на 1—2 °С. П. інактівіруєт багато гормонів: тіроксин, естроген, гонадотропні гормони, стероїди кори надниркових, серотонін і ін. Деякі речовини, пройшовши через П., набувають великої токсичності, наприклад алкалоїд колхіцин перетворюється на отруйнішу речовину — оксиколхицин; сульфаніламіди після ацетилювання в П. стають менш розчинними, унаслідок чого осідають в сечових дорогах. П. володіє желчеобразовательной функцією. Жовч, що синтезується в клітках П. з речовин, що поступають з кров'ю, має важливе значення для процесів травлення і жирового обміну в організмі. Не менш важлива функція П.— депонування крові (див. Депо крові ). Судини П. можуть вміщати 20% всій крові, тому затримка крові в нормальній здоровій П. не є венозним застоєм. Від її функціонального стану залежить діяльність ін. органів, що депонують кров (селезінки, кишечника). Вся кров, що вийшла з селезінки і кишечника, обов'язково проходіт через П. Здесь віддаляється надлишок води з крові, який йде на створення лімфа і жовч. У П. утворюється від 1 / 3 до 1 / 2 всієї лімфи з великим вмістом білка (6%).
До складу П. входять: вода (70—75%), прості і складні білки (12—24%) і продукти їх розпаду, ліпіди (2—6%), вуглеводи (2—8%) і продукти їх розщеплювання, коферменти, вітаміни, гормони, всілякі низькомолекулярні органічні речовини і мінеральні катіони і аніони. П. виконує вельми важливі функції. У їй проходят процеси біосинтезу найважливіших для організму з'єднань — нуклеїнових кислот (ДНК і РНК(рибонуклеїнова кислота)), різних ді- і мононуклеотидів, пурінових і пірімідінових підстав. В той же час ферменти, що містяться в П., викликають розщеплювання нуклеїнових кислот і нуклеотидів, дезамінування і окислення вільних пурінових підстав. П. бере участь в тій або іншій мірі в обміні білків вуглеводів, ліпідів, вітамінів, мінеральних речовин і води. Продукти розщеплювання всіх живильних речовин утворюють в П. основний «метаболічний фонд», з якого організм черпає у міру потреби необхідні для нього речовини.
Білковий обмін. З 80—100 г білка, розщеплюваної людини, що знов синтезується в організмі, за добу, приблизно половина доводиться на П. Белки в П. оновлюються за 7 діб, а в ін. органах — за 17 і більш. Це свідчить про інтенсивність білкового обміну в П. В ній відбувається синтез білків, починаючи з активації амінокислот в гіалоплазмі, утворення з'єднань із специфічними для кожної амінокислоти транспортними РНК(рибонуклеїнова кислота) і кінчаючи завершуючою стадією синтезу — вивільненням довгих пептидних ланцюгів готових білків з місця їх синтезу в рибосомах. У П. утворюються не лише білки, характерні для неї самої, але і білки плазми крові — альбумін, багато глобуліни, а також фібриноген і ін. чинники, що беруть участь в процесі згортання крові . Під впливом катептічеських протеаз і пептидази (див. Катепсини .) в П. відбувається розщеплювання білків і утворення амінокислот, які піддаються в ній різним перетворенням: дезамінуванню (практично відбувається лише в П.), переамінуванню декарбоксилюванню, що приводить до виникнення біогенних амінів; в результаті перенесення метільной групи від аденозілметіоніна забезпечується утворення холіну, креатину, адреналіну і ін. метилованих з'єднань. Своєрідні і характерні дорозі перетворення в П. окремих амінокислот — триптофану, фенілаланіна, гістидину, лізину і ін. З триптофану, зокрема, синтезуються такі біологічно активні речовини, як тріптамін, окситріптофан і продукт його декарбоксилювання — серотонін, хиноліновая кислота і обидва продукти її декарбоксилювання — нікотинова і піколінова кислоти; з гістидину утворюються форміміноглутаміновая і глутамінова кислоти, а також гістамін ; з аргініну утворюються орнітин і сечовина. Орнітин вступає в характерний цикл реакцій, що приводять до синтезу кінцевого продукту обміну простих білків — сечовини (з вуглекислого газу і аміаку за участю іонів магнію, АТФ і ряду амінокислот — див.(дивися) Орнітіновий цикл ). У П. відбуваються синтези, нейтралізуючі токсичні продукти обміну речовин, такі, як феноли, ароматичні вуглеводні і багато ін. з утворенням гиппурової і фенацетурової кислот (при використанні гліцину), а також парних глюкуронових ефіросерних кислот меркаптокислот і ін. з'єднань.
Вуглеводний обмін. П. підтримує концентрацію цукру в крові на такому рівні, який забезпечує безперервне постачання глюкозою всіх тканин. Це досягається регуляцією співвідношення між синтезом і розпадом глікогену, що депонується в П. (див. Кори цикл ). У середньому П. людини містить 30—100 г глікогену. Цієї кількості вистачає, щоб служити резервуаром для регуляції рівня цукру в крові. При всмоктуванні цукру з кишечника вміст глюкози в крові комірної вени може підвищуватися до 400 мг% , а в периферичній крові її міститься не більше 200 мг% Глюкоза перетворюється на П. в глікоген і депонується, а також використовується для здобуття енергії. Якщо після цього і ін. синтезів ще є надлишок глюкози, вона перетворюється на жир. При голодуванні П. підтримує постійний рівень цукру в крові перш за все розщеплюванням глікогену; якщо цього недостатньо — гліконеогенезом (перетворенням глікогенних амінокислот і гліцерину на цукор). Інсулін, що утворюється у внутрішньосекреторних відділах підшлункової залози, проходячи через П., також робить вплив на рівень цукру в крові і на освіту і розпад глікогену в П. Под впливом фосфорилази кінцеві глюкозниє залишки глікогену відщеплюються з утворенням глюкозо-1-фосфата, що бере участь в утворенні урідіндіфосфатглюкози, — транспортної форми глюкозних залишків і основного їх джерела при синтезі глікогену. Порушення ферментативного перетворення галактозо-1-фосфата в глюкозо-1-фосфат приводить до важких патологічних явищ, пов'язаних із спадковою хворобою, — галактоземієй. Звичайна дорога перетворення глюкозо-1-фосфата (освіта з нього глюкозо-6-фосфата) має велике біологічне значення, оскільки це з'єднання грає центральну роль в перетвореннях вуглеводів і саморегуляції вуглеводного обміну . У П. глюкозо-6-фосфат різко гальмує фосфоролітічеськоє розщеплювання глікогену, активує ферментативний транспорт глюкози з урідінфосфоглюкози на молекулу глікогену, що будується, є субстратом для окислювального перетворення глюкози по пентозофосфатному дорозі. При окисленні глюкозо-6-фосфата утворюється відновлена форма никотінамідаденіндінуклеотідфосфата (НАДФ) — необхідного кофермента відновних синтезів жирних кислот і холестерину і перетворення глюкозо-6-фосфата у фосфопентози — обов'язковий компонент при утворенні нуклеотидів і нуклеїнових кислот. Крім того, глюкозо-6-фосфат — субстрат для подальших гліколітичних перетворень, що приводять через фруктозомоно- і діфосфати до фосфотріозам і утворення піровиноградної і молочної кислот. Цей процес забезпечує організм з'єднаннями, необхідними для біосинтезів, і грає істотну роль в обміні енергії, оскільки утворення кожної молекули молочної кислоти рівноцінне синтезу однією багатою енергією фосфатного зв'язку в молекулі АТФ. Нарешті, розщеплювання глюкозо-6-фосфата фосфатазою забезпечує вступ в кров вільної глюкози, крові, що доставляється струмом, в всі органи і тканини.
Жировий обмін. П. в змозі депонувати значно більше ліпідів, чим глікогену (до 20—30% сухої маси). Частина ліпідів, що складається з фосфатідов і холестерину, досить постійна і складає 10—15%; вміст нейтрального жиру вагається. Депонування жиру — функція жирової тканини, а не П. В цілому П. не грає в ліпідному відміні такої життєво важливої ролі, як у вуглеводному і білковому обмінах. Розщеплювання жирних кислот також не обмежується П. В П. відбуваються деградація жиру і окислення жирних кислот, а також представлені ферментні системи біосинтезу високомолекулярних жирних кислот, нейтрального жиру і складних ліпідів; проміжний продукт при цих синтезах — фосфатідная кислота. У П. синтезується також холестерин. Що утворюються при деградації жиру жирні кислоти окислюються з утворенням ацетилкофермента A, вступаючого за наявності конденсуючого ферменту в реакцію з щавлевооцетовою кислотою і створюючого таким чином лимонну кислоту — основний субстрат окислювальних перетворень в трікарбонових кислот циклі . У клітках П., як і в клітках ін. органів, окислювального перетворення, локалізовані по перевазі в мітохондріях, зв'язані з утворенням багатих енергією з'єднань (АТФ) і закінчуються утворенням Co 2 і H 2 O. Синтез високомолекулярних жирних кислот протікає поза мітохондріями — в так званому цитозоле і, отже, просторово відокремлений від місця їх окислення. У так званій мікросомальной фракції П. зосереджена 2-я (не мітохондріальна) НАДФ-залежна система окислення вуглеводнів, стероїдів, холестерину. Ця система локалізована в ендоплазматичному ретікулуме і пов'язана з утворенням продуктів гідроксилювання. П. має істотне значення в про міну пігментів: у ній руйнується гемоглобін, утворюється білірубін і перетворюється на розчинну форму у вигляді діглюкуроніда білірубіну. Пігментний обмін в П., тісно пов'язаний з метаболізмом білірубіну і порфірінов, у свою чергу, грає важливу роль в обміні заліза в організмі.
В мінеральному обміні і в збереженні постійності кислотно-лужного рівноваги П. бере безпосередню участь. Мінеральні речовини в П. знаходяться як у вільному вигляді, так і входять до складу складних органічних сполук, наприклад ферментів (Mg, Mn, Fe, Cu, Zn). Катіони виконують також роль активаторів ферментів, наприклад Na + , Ca 2+ , К + , Ni 2- , Co 2+ , Cr 3+ і ін. У складі П. знаходяться залізовмісний білок ферритин і медьсодержащий білок гепатокупрєїн; ці речовини беруть участь в процесі кровотворення . П. також бере участь в обміні вітамінів. У ній містяться вітаміни групи В і D, вітамін C і розчинні в жирах вітаміни E і K. З каротинів в П. утворюється і депонується вітамін A, всмоктування якого з кишечника відбувається лише у присутності жовчі. Аскорбінова кислота сприяє глікогенезу в П. Вітамін До необхідний для синтезу протромбіну в ній.
Функції П. (процеси обміну речовин, що протікають в ній, жовчоутворення) регулюються нервовими і гормональними механізмами. У гормональній регуляції беруть участь адреналін, інсулін, глюкагон, кортикостероїди, гормони, що виробляються в гіпофізі, інтестінальниє гормони, особливо секретин, холецистокінін, панкреозимін. Вплив багатьох гормональних чинників реалізується в П. при участі циклічних мононуклеотидів: циклічного аденозінмонофосфата (цАМФ) і гуанозінмонофосфата (цГМФ). Ці циклічні мононуклеотиди утворюються при розщеплюванні циклазой (ферментом, фіксованим головним чином в плазматичній мембрані) нуклеозідтріфосфатов АТФ і ГТф. Циклічні мононуклеотиди виконують функції регулювальників активності багатьох ферментів в результаті активації протєїнкиназ, що забезпечують процес перенесення фосфатного залишку з АТФ на білки-ферменти. Фосфорилування ферментів міняє їх активність, підвищуючи її в одних (фосфорилази, ліпази) і пригнічуючи в інших (глікоген-синтетази, піруватдекарбоксилази).
У свою чергу, біохімічні процеси, що протікають в П., через тканинні інтероцептори роблять вплив на функціональний стан центральної нервової системи. Різноманітність і взаємозалежність чинників, що впливають на стан кліток П., обумовлюють як інтенсивність, так і спрямованість процесів обміну речовин, що протікають в ній.
При захворюваннях П. у людини і тварин приголомшується переважно її паренхіма (клітки) або проміжна тканина. Гострі гепатити складають значну частину всіх хвороб П. і можуть бути причиною розвитку хронічних її поразок. Серед гепатитів інфекційної природи розрізняють первинні (див. Гепатит вірусний ) і вторинні (наприклад, при бруцельозі, лептоспірозі, сифілісі і ін.). Токсико-алергічні гепатити розвиваються при дії на організм хімічних, у тому числі лікарських речовин (див. Алергія ). Результатом хвороби може бути цироз печінки . В результаті порушення живлення і обміну речовин (при алкоголізмі, вітамінній недостатності, патологічному голодуванні, діабеті цукровому, гепато-церебральній дистрофії, ожирінні і т.д.), а також при порушеннях кровообігу, особливо при підвищенні тиску в ніжней порожнистій і печінковій венах, в П. розвиваються дистрофічні зміни. П. людини і тварин може бути местомом проживання багатьох паразитів: простих гельмінтів, рідше членистоногих. Так, з кишечника в жовчні дороги можуть проникати лямблії, лейшманії (див. Лямбліоз, Лейшманіоз ); у П. людини відбувається позаеритроцитарний розвиток збудника малярії . У протоках П. і жовчному міхурі паразитують різні трематоди, що викликають холангіти, холецистити (див. Дікроцеліоз, Клонорхоз, Опісторхоз, Шистосоматози, Фасциолез ). П. може бути местомом розвитку збудників ехінококозу . З новоутворень П. зустрічаються рак, саркома і ін.
Літ.: Догель Ст А., Порівняльна анатомія безхребетних, ч. 1, Л., 1938; Шмальгаузен І. І., Основи порівняльній анатомії хребетних тварин, 4 видавництва, М., 1947; Павлов І. П., Лекції з фізіології, Полн. собр. соч.(вигадування), 2 видавництва, т. 5, М.— Л., 1952; Фішер А., Фізіологія і експериментальна патологія печінки, пер.(переведення) з англ.(англійський), Будапешт, 1961; Рапопорт С. М., Медична біохімія, пер.(переведення) з йому.(німецький), М., 1966; Введення в клінічну біохімію (основи патобіохимії), Л., 1969: Бондар З. А., Клінічна гепатологія, М., 1970; Блюгер А. Ф., Райцис А. Б., Серотонін і печінка, «Успіхи гепатології», ст 3, Рига, 1971; Збарський Би. І., Іванов І. І., Мардашев С. Р., Біологічна хімія, 5 видавництво, Л., 1972.