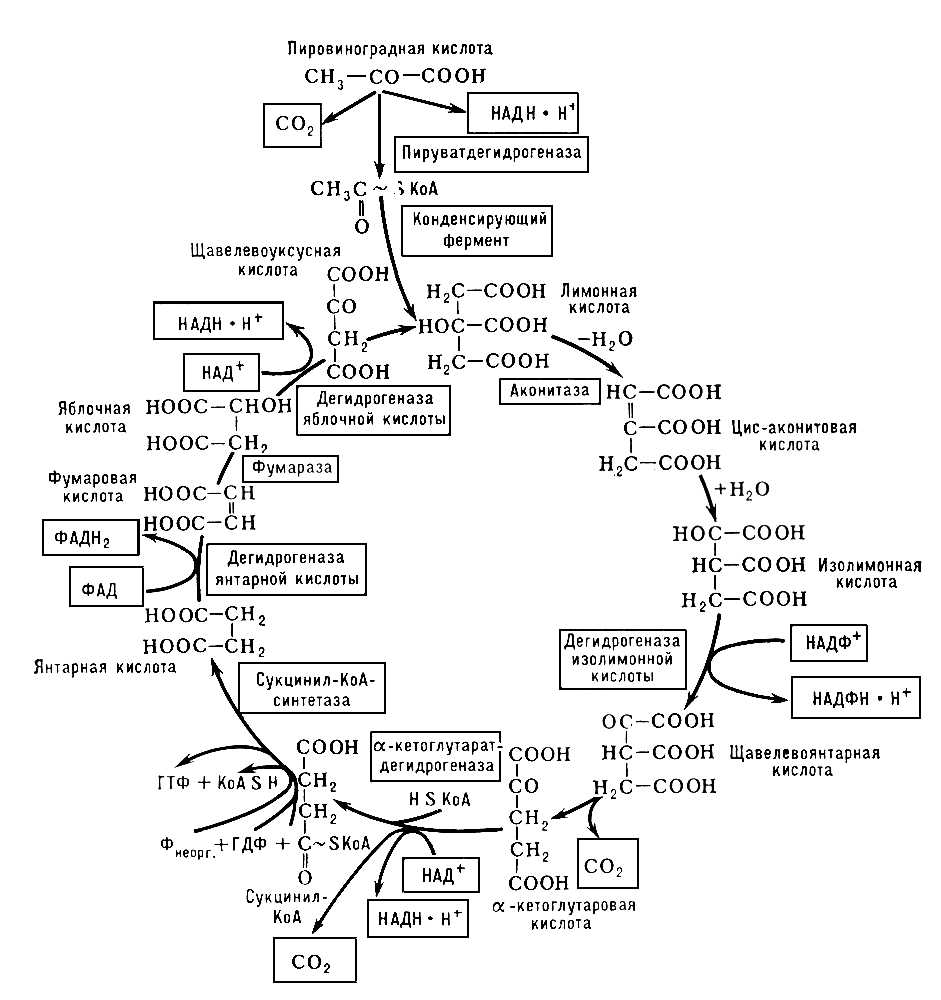
Трікарбонових кислот цикл , цикл лимонної кислоти, цикл Кребса, широко представлений в організмах тварин, рослин і мікробів дорога окислювальних перетворень ді- і трікарбонових кислот, що утворюються як проміжні продукти при розпаді білків, жирів і вуглеводів. Відкритий Х. Кребсом і У. Джонсоном (1937). Т. до. ц., локалізований в мітохондріях, починається з лимонної кислоти і закінчується утворенням щавлевооцетової кислоти, Co 2 і відновленням коферментів дегідрогенази: никотінамідаденіндінуклеотіда (НАД) і флавінаденіндінуклеотіда (ФАД). До субстратів Т. до. ц. відносяться трікарбоновиє кислоти — лимонна, цис-аконітова, ізолимонна, щавельовоянтарная і дикарбонові кислоти — кетоглутарова, янтарна, фумарова, яблучна і щавлевооцетова. До субстратів Т. до. ц. слід віднести і оцетову кислоту, яка в активній формі, тобто у вигляді ацетилкофермента А, бере участь в конденсації з щавлевооцетовою кислотою, що приводить до утворення лимонної кислоти. Саме ацетиловий залишок, що увійшов до структури лимонної кислоти, піддається окисленню; атоми вуглецю окислюються до Co 2 , атоми водню частково акцептуються коферментами дегідрогенази, частково в протонованій формі переходять в розчин, тобто в довкілля.
Зазвичай вказують на піровиноградну кислоту (піруват), що утворюється при гліколізі в реакціях переамінування і що займає одне з центральних місць в дорогах обміну речовин, що перехрещуються, як на вихідне з'єднання для утворення ацетіл-КоА. Дійсно, під впливом ферменту складної структури — піруватдегидрогенази — здійснюється окислення пірувату з утворенням Co 2 (перше декарбоксилювання), ацетіл-КоА і відбувається відновлення НАД (див. схему ). Проте окислення пірувату далеке не єдина дорога утворення ацетіл-КоА, який є характерним продуктом мітохондріального окислення жирних кислот (фермент тіолаза), а також реакції зворотної конденсації при утворенні лимонної кислоти і ін. Всі ферменти, що беруть участь в реакціях Т. до. ц., локалізовані в мітохондріях, причому більшість з них міцно пов'язані з мембранними структурами.
Утворення лимонної кислоти, з перетворення якої і починається власне Т. до. ц., є реакцією ендергонічеськой, і її реалізація можлива завдяки використанню багатою енергією зв'язку ацетилового остагка з KOA [СН 3 (О) С~skoa]. Далі слідує ізомеризація лимонної кислоти в ізолимонну через проміжну стадію утворення цис-аконітової кислоти. Продуктом подальшого перетворення ізолимонної кислоти під впливом відповідної дегідрогенази є, мабуть, щавельовоянтарная кислота, декарбоксилювання якої (друга молекула Co 2 ) приводить до а-кетоглутарової кислоти. Кетоглутаратдегидрогеназа по ряду характеристик (висока молекулярна маса, складна багатокомпонентна структура, ступінчасті реакції, частково ті ж коферменти і так далі) нагадує дію піруватдегидрогенази. Продуктами реакції є Co 2 (третє декарбоксилювання), НАДН×Н + і сукциніл-КоА. На цій стадії включається сукциніл-КоА-синтетаза, що каталізує оборотну реакцію утворення вільного сукцинату: Сукциніл-КоА + Р неорг. + ГДФ Û Сукцинат + KOA + ГТФ. При цій реакції здійснюється так зване субстратноє фосфорилування, тобто освіта багатого енергією гуанозінтріфосфата (ГТФ) або аденозинтрифосфату (АТФ) за рахунок гуанозіндіфосфата (ГДФ) і мінерального фосфату (Р) з використанням енергії сукциніл-КоА. Після утворення сукцинату вступає в дію сукцинатдегидрогеназа — флавопротеїд, що приводить до фумаровій кислоті. Фумараза забезпечує рівновагу між фумаровою кислотою і яблучною, а дегідрогеназа яблучної кислоти (кофермент — НАД+) приводить до завершення Т. до. ц., тобто до утворення щавлевооцетової кислоти. На цій стадії повторюється реакція конденсації (конденсуючий фермент) між щавлевооцетовою кислотою і ацетіл-КоА, що приводить до утворення лимонної кислоти.
Енергетична ефективність розглянутих процесів невелика. Що проте утворюються при окисленні пірувату і подальших реакціях Т. до. ц. 4 благаючи НАДН, 1 міль ФАДН 2 і 3 благаючи Co 2 є важливими продуктами окислювальних перетворень. Особливо це стосується відновлених форм НАД і ФАД. Подальше їх окислення здійснюється ферментами дихального ланцюга і зв'язано з фосфорилуванням, тобто освітою АТФ за рахунок етерифікації мінерального фосфату (див. Окислювальне фосфорилування ). На кожну повністю окислену до Co 2 і H 2 O молекулу пірувату доводиться освіта не менше 15 багатих енергією фосфатних зв'язків. Процес окислення НАДН і ФАДН 2 ферментами дихального ланцюга енергетично вельми ефективний, відбувається з використанням кисню повітря, приводить до утворення води і служить основним джерелом енергетичних ресурсів клітки. Проте в його безпосередній реалізації ферменти Т. до. ц. не беруть участь. Див. також Окислення біологічне, Тканинне дихання .
Літ.: Кребс Р., Корнберг Р., Перетворення енергії в живих системах, пер.(переведення) з англ.(англійський), М., 1959; Пилипович Ю. Б., Основи біохімії, М., 1969; Ленінджер А., Біохімія, пер.(переведення) з англ.(англійський), М., 1974, гл.(глав) 16.