Окислення біологічне , сукупність реакцій окислення, що протікають у всіх живих клітинах. Основна функція О. б. — забезпечення організму енергією в доступній для використання формі. Реакції О. б. у клітках каталізують ферменти, що об'єднуються в клас оксидоредуктази . Вивчення окислення в організмі було почате в 18 ст А. Лавуазье ; надалі значний вклад в дослідження О. б. (його локалізація в живих клітинах, зв'язок з ін. процесами обміну речовин, механізми ферментативних окислювально-відновних реакцій, акумуляція і перетворення енергії і ін.) внесли О. Варбург, Р. Віланд (Німеччина), Д. Кейлін, Х. Кребс, П. Мітчелл (Великобританія), Д. Грін, А. Ленінджер, Би. Чанс, Е. Рекер (США), а в СРСР — А. Н. Бах, Ст І. Палладін, В . А. Енгельгардт, С. Е. Северин, Ст А. Беліцер, В. П. Ськулачев і ін.
О. б. у клітках пов'язано з передачею т. н. поновлюючих еквівалентів (ВЕ) — атомів водню або електронів — від одного з'єднання — донора, до іншого — акцептору. В аеробів — більшості тварин, рослин і багатьох мікроорганізмів — кінцевим акцептором ВЕ служить кисень. Постачальниками ВЕ можуть бути як органічні, так і неорганічні речовини (див. таблицю).
Класифікація організмів за джерелом енергії і поновлюючих еквівалентів
Тип організмів
Джерело енергії
Окислюване з'єднання (постачальник поновлюючих еквівалентів)
Приклади
Фотолітотрофи
Фотоорганотрофи
Хемолітотрофи
Хемоорганотрофи
Світло
Світло
Реакції окислення
Реакції окислення
Неорганічні з'єднання (Н 2 Про, H 2 S, S)
Органічні сполуки
Неорганічні з'єднання (H 2 , S, H 2 S, Nh 3 , Fe 2 + )
Органічні сполуки
Зелені клітки вищих рослин, синезелениє водорості, фотосинтезуючі бактерії
Тварини, більшість мікроорганізмів, нефотосинтезуючі клітки рослин
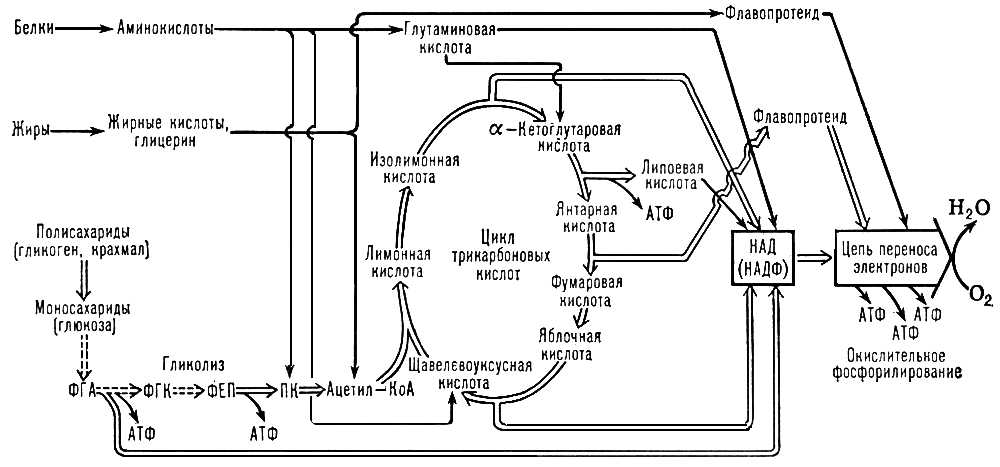
Основна дорога використання енергії, що звільняється при О. б., — накопичення її в молекулах аденозинтрифосфорної кислоти (АТФ) і ін. макроергічних з'єднань . О. б., що супроводиться синтезом АТФ з аденозіндіфосфорной кислоти (АДФ) і неорганічного фосфату, відбувається при гліколізі, окисленні а-кетоглутарової кислоти і при перенесенні ВЕ в ланцюзі окислювальних (дихательних) ферментів, зазвичай називають окислювальним фосфорилуванням (см. схему ).
В процесі дихання вуглеводи, жири і білки піддаються багатоступінчастому окисленню, яке приводить до відновлення основних постачальників ВЕ для дихальних флавінов, никотінамідаденіндінуклеотіда (НАД), никотінамідаденіндінуклеотідфосфата (НАДФ) і ліпоєвої кислоти . Відновлення цих з'єднань значною мірою здійснюється в трікарбонових кислот циклі, яким завершуються основні дороги окислювального розщеплювання вуглеводів (воно починається з гліколізу), жирів і амінокислот. Окрім циклу трікарбонових кислот, деяка кількість відновлених коферментів — ФАД (флавінаденіндінуклеотіда ) і НАД — утворюється при окисленні жирних кислот, а також при окислювальному дезамінуванні глутамінової кислоти (НАД) і в пентозофосфатном циклі (відновлений НАДФ).
Співвідношення і локалізація різних механізмів О. б. З розрахунку на 1 молекулу глюкози гліколіз дає 2 молекули АТФ, а фосфорилування в дихальному ланцюзі — 34 молекули АТФ. Гліколіз, цикл трікарбонових кислот і дихальний ланцюг функціонують, мабуть, в клітках всіх еукаріотов . Окислення жирних кислот в хребетних поставляє половину енергії, споживаною печінкою, нирками, м'язом серця і скелетними м'язами, що покояться; у клітинах мозку воно практично не відбувається. Окислення по пентозофосфатному дороги активно в печінці і лактірующих молочних залозах, але трохи в сердечній і скелетних м'язах.
В рідкій фазі цитоплазми розчинені всі ферменти гліколізу. Внутрішні мембрани мітохондрій, мембрани хлоропластів (тілакоїдов) і клітинні мембрани бактерій містять ланцюги перенесення, що фосфорилюють електронів. У матриксі мітохондрій локалізовано окислення жирних кислот, ферменти циклу трікарбонових кислот і глутаматдегидрогеназа. У внутрішній мембрані мітохондрій знаходяться ферменти, що окислюють янтарну і b-оксимасляну кислоти, в зовнішній — ферменти, що беруть участь в обміні амінокислот: моноаміноксидаза і кинуренінгидроксилаза. У особливих органоїдах клітки, т.з. пероксисомах, або мікротельцях, вклад яких в сумарне поглинання О 2 може досягати в печінці 20%, знаходиться флавіновая оксидаза, що окислює амінокислоти, гліколеву кислоту і ін. субстрати з утворенням перекису водню, який потім розкладається каталазой або використовується пероксидазамі в реакціях окислення. У мембранах ендоплазматичної мережі клітки локалізовані гидроксилази і оксигенази, організовані в короткі ланцюги перенесення електронів, що не фосфорилюють.
Окислювальні реакції не завжди супроводяться накопиченням енергії; у ряді випадків вони несуть функції перетворення речовин (наприклад, окислення при освіті жовчних кислот, стероїдних гормонів, на дорогах перетворення амінокислот і ін.). При окисленні відбувається знешкодження чужорідних і отруйних для організму речовин (ароматичних з'єднань недоокисленних продуктів дихання і ін.). О. б., не зв'язане з накопиченням енергії, називається вільним окисленням. Його енергетичний ефект — утворення тепла. Мабуть, система перенесення електронів, що здійснює окислювальне фосфорилування, здатна перемикатися на вільне окислення при збільшенні потреби організму в теплі (в гомойотермних тварин ).
Механізм використання енергії окислення . Довгий час залишалося неясним питання про механізм перетворення енергії, що звільняється при перенесенні ВЕ по ланцюгу окислювальних ферментів. Згідно т.з. хеміосмотічеськой теорії, розвиненій в 60-х рр. 20 ст (англійський біохімік П. Мітчелл і ін.), енергія спочатку використовується для створення електричного поля («+» з одного боку мембрани і «–» з іншою) і різниці концентрацій іонів Н + по різні сторони мембрани. Обидва чинники (електричне поле і різниця концентрацій) можуть служити рушійною силою для дії ферменту АТФ-СИНТЕТАЗИ, що здійснює синтез АТФ. Частина енергії поля може бути прямо використана кліткою для перенесення іонів через мембрану, відновлення переносників електронів, утворення тепла без проміжної участі АТФ.
Еволюція енергозабезпечення в живій природі . Прадавні організми, як вважають, існували в первинній безкисневій атмосфері Землі і були анаеробами і гетеротрофними організмами . Забезпечення кліток енергією йшло за рахунок процесів типа гліколізу. Можливо, існував механізм окислення, відомий у деяких сучасних мікроорганізмів: ВЕ передаються через дихательную ланцюг на нітрат (No – 3 ) або на сульфат (So – 4 ). Принципово важливим етапом виявилося виникнення в древніх одноклітинних організмів механізму фотосинтезу, з яким зв'язують появу кисню в атмосфері Землі. В результаті стало можливим використання O 2 , що володіє високим окислювально-відновним потенціалом, як кінцевий акцептор електронів в дихальному ланцюзі. Реалізація цій можливості сталася при появі спеціального ферменту — цитохромоксидази, поновлюючою О 2 , і привела до виникнення біохімічного дихального апарату сучасного типа. Забезпечення енергією у всіх аеробів (їх клітки містять мітохондрії) засноване на такому диханні. В той же час клітки зберегли ферментний апарат гліколізу. Утворювана в ході останнього піровиноградна кислота окислюється далі в циклі трікарбонових кислот, який, у свою чергу, живить дихальний ланцюг електронами. Т. о., еволюція енергетичного обміну йшла, мабуть, по шляху використання і надбудови механізмів енергозабезпечення, що вже були раніше. Наявність в клітках нині існуючих організмів біохімічних систем гліколізу (у цитоплазмі), дихання (у мітохондріях), фотосинтезу (у хлоропластах), а також вражаюча схожість механізмів перетворення енергії в цих органелах і в мікроорганізмах незрідка розглядають як свідоцтво можливого походження хлоропластів і мітохондрій від древніх мікроорганізмов-симбіонтов. Див. також Аденозінфосфорниє кислоти,Біоенергетика,Бродіння,Дихання,Мітохондрії,Фотосинтез і літ.(літературний) при цих статтях.
Літ.: Ленінгер А., Перетворення енергії в клітці, в кн.: Жива клітина, пер.(переведення) з англ.(англійський), М., 1962; Ськулачев Ст П., Акумуляція енергії в клітці, М., 1969; його ж, Трансформація енергії в біомембранах, М., 1972; Малер Г. і Кордес Ю., Основи біологічної хімії, пер.(переведення) з англ.(англійський), М., 1970, гл.(глав) 15; Льові А., Сикевіц Ф., Структура і функції клітки, пер.(переведення) з англ.(англійський), М., 1971, гл.(глав) 12; Ясайтіс А. А., Перетворення енергії в мітохондріях, М., 1973; Ленінджер А., Біохімія, пер.(переведення) з англ.(англійський), М., 1974.