Фотосинтез (від фото... і синтез ) , освіта вищими рослинами, водоростями, фотосинтезуючими бактеріями складних органічних речовин, необхідних для життєдіяльності як самих рослин, так і всіх ін. організмів, з простих з'єднань (наприклад, вуглекислого газу і води) за рахунок енергії світла, що поглинається хлорофілом і ін. фотосинтетичними пігментами. Один з найважливіших біологічних процесів, постійно і у величезних масштабах що здійснюється на нашій планеті. В результаті Ф. рослинність земної кулі щорік утворює більше 100 млрд. т органічного речовин (близько половини цієї кількості припадає на частку Ф. рослин Морея і океанів), засвоюючи при цьому близько 200 млрд. т Co 2 і виділяючи в зовнішнє середовище близько 145 млрд. т вільного кисню. Вважають, що завдяки Ф. утворюється весь кисень атмосфери. Ф. – єдиний біологічний процес, який йде із збільшенням вільної енергії системи; всі інші (за винятком хемосинтезу ) здійснюються за рахунок потенційної енергії, що запасається в продуктах Ф. Колічество енергії, що щорік зв'язується фотосинтезуючими організмами океану і суші (біля 3×10 21 дж ), у багато разів більше тієї енергії, яка використовується людством (біля 3×10 20 дж ) .
Історична довідка. Почало дослідженню Ф. належить роботами Дж. Прістлі, Же. Сенебье, Н. Соссюра, Я. Інгенхауза, Ю. Майєру, в яких поступово з'ясувалося, що рослини на світлу засвоюють з повітря вуглекислий газ, виділяють кисень, утворюють в результаті цього органічні речовини, запасаючи в них енергію сонячного світла. У 2-ій половині 19 ст До. А. Тімірязев показав що енергія сонячного світла вводиться в ланцюг фотосинтетичних перетворень через зелений пігмент рослин – хлорофіл : спектр дії Ф. відповідає спектру поглинання світла хлорофілом, і інтенсивність Ф. збільшується із збільшенням інтенсивності світла. У 1905 англійським учений Ф. Блекман виявив, що Ф. складається з швидкої світлової реакції і повільнішою – темнової. Біохімічний доказ існування світлової і темнової фаз було отримане лише в 1937 англійським дослідником Р. Хиллом. Крупний вклад у вивчення темнової і світлових стадій Ф. внесли також йому.(німецький) біохімік і фізіолог О. Варбург, амер.(американський) біохімік Х. Гафрон . В 1931 амер.(американський) мікробіолог До. Нил показав, що фототрофниє бактерії здійснюють Ф. без виділення О 2 , т.к. прі асиміляцію СО 2 окислюють сірководень, тіосульфат і ін. субстрати. Так належало початок уявленню про Ф. як окислювально-відновному процесі, де відновлення Co 2 здійснюється при одночасному окисленні донора водню. У 1941 сов.(радянський) ученими А. П. Віноградовим і М. Ст Тейц, а також американськими дослідниками Е. Рубеном і ін. встановлено, що джерелом кисню, що виділяється в процесі Ф. вищих рослин і водоростей, є вода, а не Co 2 , як вважали раніше.
Починаючи з 1-ої чверті 20 ст важливі роботи виконані по вивченню фізіології і екології Ф. (Ст Ст Чоботарів, С. П. Костичев, Ст Н. Любіменко, А. А. Нічипоровіч, О. В. Заленський і багато ін.). З середини 20 ст вивченню Ф. сприяло створення нових методів дослідження (газовий аналіз, ізотопні методи, спектроскопія, електронна мікроскопія і ін.). Ці методи дозволили розробити уявлення про тонкі механізми участі хлорофілу у Ф. (А. Н. Теренін, А. А. Красновський, американські учені Е. Рабіновіч, Ст Кок, В. Арнолд, Р. Клейтон, Дж. Франк, франц.(французький) дослідник Дж. Лаворель); про окислювально-відновні реакції Ф. і про існування двох фотохімічних реакцій Ф. (англійський фітофізіолог Р. Хилл, С. Очоа, амер.(американський) дослідники Ст вишневий Мед, Р. Емерсон, Френч, голландський вчений Л. Дейсенс); про фотосинтетичне фосфорилування (Д. Арнон ) ; про дороги перетворення вуглецю (М. Калвін, амер.(американський) учені Дж. Бассам, Е. Бенсон, австралійські дослідники М. Хетч і К. Слек); про механізм розкладання води (Ст Кок, французькі учені А. і П. Жоліо, радянський учений В. М. Кутюрін і ін.).
Характерні межі фотосинтезу вищих зелених рослин, водоростей і ф отосинтезірующих бактерій. В реакціях Ф. у вищих зелених рослин, водоростей (багатоклітинних – зелених, бурих, червоних, а також одноклітинних – евгленових, дінофлагеллят, діатомових) донором водню і джерелом кисню, що виділяється, служить вода, а основним акцептором атомів водню і джерелом вуглецю – вуглекислий газ.(газета) При використанні у Ф. лише Co 2 і H 2 O утворюються вуглеводи. Але в процесі Ф. рослини утворюють не лише вуглеводи, але і азот, що містять, і сірку амінокислоти, білки, а також пігменти і ін. з'єднання. Акцепторами атомів водню (поряд з Co 2 ) і джерелами азоту і сірки в цьому випадку служать нітрати () і сульфати (). Фотосинтезуючі бактерії не виділяють і не використовують молекулярний кисень (більшість з них облігатні, тобто обов'язкові анаероби ) . Замість води як донорів електронів ці бактерії використовують або неорганічні з'єднання (сірководень, тіосульфат, газоподібний водень), або органічні речовини (молочну кислоту, ізопропіловий спирт). Джерелом вуглецю в більшості випадків є також Co 2 , але поряд з цим і деякі органічні сполуки (наприклад, ацетат). Т. о., Ф. в різних організмів може протікати з використанням різних донорів (ДН 2 ), акцепторів (А) електронів і водню і може бути представлений схематично узагальненим рівнянням:
Д×Н 2 + А Ah 2 + Д,
де Ah 2 – продукти Ф.
Структурні особливості фотосинтетичного апарату. Висока ефективність Ф. вищих зелених рослин забезпечується досконалим фотосинтетичним апаратом, основа якого – внутріклітинні органели – хлоропласти (у клітці зеленого аркуша їх 20–100). Вони оточені двуслойной мембраною. Внутрішній шар її побудований з мішечків сплощень або бульбашок, званих тілакоїдамі, які часто упаковані в стопки, складають грани, сполучені між собою одіночнимі межграннимі тілакоїдамі. Тілакоїди складаються з власне фотосинтетичних мембран, що є біомолекулярними ліпідними шарами і мозаїчно вкраплені в них ліпопротєїдопігментниє комплекси, створюючі фотохімічно активні центри, і містять також спеціальні компоненти, що беруть участь в транспорті електронів і утворенні аденозинтрифосфату (АТФ). Частина хлоропласту, що знаходиться між тілакоїдамі строма містить ферменти, що каталізують темновиє реакції Ф. (наприклад, перетворення вуглецю, азоту, сірки, біосинтез вуглеводів і білків). У стромі відкладається утворюваний при Ф. крохмаль. Хлоропласти має власна ДНК(дезоксирибонуклеїнова кислота), РНК(рибонуклеїнова кислота), рибосоми, синтезуючі білки, і володіють деякою генетичною автономністю, але знаходяться під загальним контролем ядра. що фотосинтезують бактерії і більшість водоростей не мають хлоропластів. Фотосинтетичний апарат більшості водоростей представлений спеціалізованими внутріклітинними органелами – хроматофорами, а фотосинтезуючих бактерій і синьо-зелених водоростей – тілакоїдамі (мембрани їх містять пігмент бактеріо-хлорофіл або бактеріовірідін, а також ін. компоненти реакцій Ф.), зануреними в периферичні шари цитоплазми.
Фаза первинних перетворень і запасання енергії в процесі Ф. У основі Ф. рослин лежить окислювально-відновний процес, в якому 4 електрони (і протона) піднімаються від рівня окислювально-відновного потенціалу, відповідного окисленню води (+ 0,8 в ) до рівня, відповідного відновленню Co 2 з утворенням вуглеводів (– 0,4 в ). При цьому збільшення вільної енергії реакції відновлення Co 2 до рівня вуглеводів складає 120 ккал/міль, а сумарне рівняння Ф. виражається як:
Co 2 + H 2 O C (H 2 O) + O 2 + 120 ккал/міль.
Енергія моля квантів (Ейнштейна) червоної частини спектру складає близько 40 ккал/міль. Т. о., для Ф., що йде відповідно до приведеним рівнянням, було б досить поглинання енергії 3 квантів на молекулу Co 2 (або на виділення молекули O 2 ). Проте в окислювально-відновній реакції від води до Co 2 мають бути перенесені 4 електрони, причому перенесення кожного з них здійснюється в ході двох послідовних фотохімічних реакцій. Тому квантова витрата за оптимальних умов складає 8–12 квантів на молекулу O 2 , а максимальна ефективність перетворення енергії червоного світла – біля 30° %. У польових умовах унаслідок неповного поглинання світла, енергетичних витрат на дихання і ін. втрат, а також обмеженості вегетаційного періоду ефективність засвоєння сонячної енергії з.-х.(сільськогосподарський) рослинами в помірних широтах зазвичай не вище 0,5–1,3%. Зіставлення цих цифр з теоретичним максимальним значенням вказує на існування значних резервів, які можуть бути використані в майбутньому. Для деяких культур з.-х.(сільськогосподарський) рослин удається в спеціальних умовах підвищити енергетичну ефективність до 5–6% і навіть вище (при культивуванні водоростей до 7–10%).
Ні Co 2 , ні вода безпосередньо не поглинають світло, посередником у взаємодії цих з'єднань з квантами служить хлорофіл а, включений в структуру хлоропласту або хроматофора і створюючий функціональні фотосинтетичні одиниці, що складаються з декількох сотень молекул пігменту і реакційних центрів. Основна частина супроводжуючих пігментів (хлорофіл b, каротиноїди, фікобіліни і ін. і короткохвильові форми хлорофілу а ) виконує функцію светособірающей антени. При поглинанні квантів їх молекули переходять в збуджений стан, який шляхом міграції енергії передається на молекулу хлорофілу а, що знаходиться в реакційному центрі. Ефективність передачі енергії обумовлена близьким розташуванням молекул, а також наявністю декількох агрегованих форм хлорофілу а, реакційних центрів, що беруть участь у формуванні, і створюючих низхідні сходи енергетичних рівнів. Можливе напівпровідникове перенесення електрона по агрегованому пігменту. У реакційному центрі відбувається основний акт Ф. – розділення зарядів з подальшим утворенням первинного окислювача і первинного відновника. Існують два типи центрів ( мал. 1 ), один з яких включений в пігментну фотосистему I (ФС I), а ін. – у фотосистему II (ФС II). У фотореакції, пов'язаній з розкладанням води, бере участь ФС II: пігментом її центру служить хлорофіл а з максимумом поглинання 680 нм, гіпотетичним первинним відновником – Q (ймовірно, цитохром ) , а первинним окислювачем – складний комплекс Z. Збудження пігментної молекули центру P 680 супроводиться розділенням зарядів і утворенням окисленого Z + , який бере участь в окисленні води і виділенні O 2 . Вважають, що в систему розкладання води, поки мало вивчену, входять невідомі ферменти, іони марганцю і бікарбонат. Первинний відновник Q (виявляється по індукції флуоресценції) ФС II передає електрон переносникам (цитохроми b, f, пластохинон пластоцианін) фотосинтетичному електронному транспортному ланцюгу до реакційного центру ФС I. Пігментом цього центру служить хлорофіл а з максимумом поглинання 700 нм, первинним відновником – неідентифіковану речовину Х. Восстановленний Х передає електрон ферредоксину – залізовмісному білку, який відновлює никотінамідаденіндінуклеотідфосфат (НАДФ). Його відновлена форма – НАДФ-Н запасає основну частину енергії світла. Ін.(Древн) частина енергії електронного потоку запасається у вигляді АТФ (фотосинтетичне фосфорилування), який утворюється на низхідній ділянці перенесення електронів між ФС II і ФС I (нециклічне фотофосфорилування) або при круговому замиканні потоку у ФС I (циклічне фотофосфорилування). Фосфорилування, можливо, відбувається по хеміосмотічеському механізму за рахунок електричного потенціалу і градієнта концентрації Н + , що виникають при індукції світлом електронного потоку у мембранних структурах тілакоїдов. Експериментально виявлено, що освітлення індукує електричний потенціал на мембрані хлоропласту. Описане послідовне з'єднання двох фотореакцій I і II найймовірніше, хоча обговорюється можливість паралельного з'єднання реакцій. Передбачають, що фотосинтезуючі бактерії здійснюють Ф. за участю лише однієї пігментної фотосистеми, проте це питання не можна вважати вирішеним. Фотофізичні і фотохімічні стадії закінчуються за 10 -12 –10 -8 сік розділенням зарядів і подальшим утворенням первинного окислювача і відновника. Кордоном первинних біофізичних і біохімічних процесів зазвичай вважають появу перших хімічно стабільних продуктів – НАДФ-Н і АТФ. Ці речовини («відновна сила») використовуються потім в темпових процесах відновлення Co 2 .
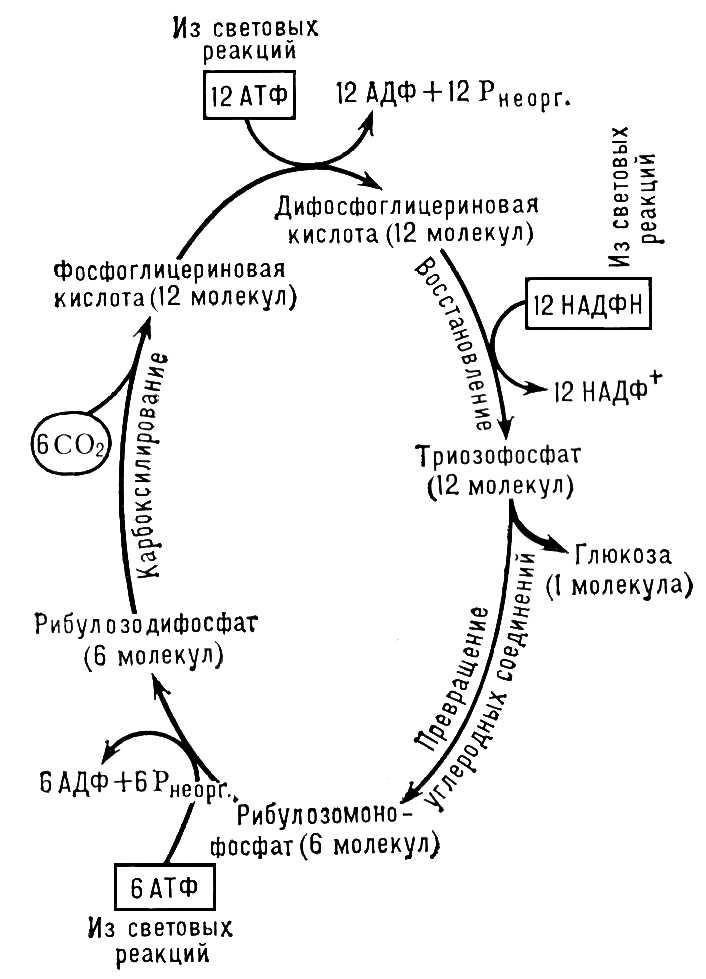
Асиміляція вуглекислоти . Асиміляція Co 2 відбувається в процесі темнових реакцій. Відновленню при Ф. піддається не вільна молекула Co 2 , а заздалегідь включена до складу певної органічної сполуки. В більшості випадків акцептором Co 2 служить двократно фосфорильований п'ятивуглецевий цукор рібулозодіфосфат (РДФ). Приєднуючи Co 2 , РДФ розпадається на 2 молекули фосфогліцерінової кислоти (ФГК). Вуглець Co 2 , включений в молекулу ФГК, і є кінцевою ланкою ланцюга, до якої прямують електрони, мобілізуємиє хлорофілом. Приєднавши електрон, ФГК перетворюється на відновлене з'єднання – фосфогліцеріновий альдегід (у цьому процесі беруть участь АТФ і НАДФ-Н), який може розглядатися як перший стабільний вуглеводний продукт Ф., вуглець, що містить, вже у відновленій (органічною) формі. Подальші перетворення відбуваються в пентозофосфатном циклі і завершуються, з одного боку, утворенням РДФ, тобто відбувається регенерація первинного акцептора Co 2 (що робить цикл за наявності світла і Co 2 що безперервно діє), а з ін. сторони – утворенням продуктів Ф. – вуглеводів.
Все, що було сказане вище, відноситься до т.з. С 3 -растеніям, які засвоюють вуглець у Ф. через цикл Калвіна ( мал. 2 ), акцептують Co 2 на РДФ при допомозі РДФ-карбоксилазі, утворюючи перші трьохвуглецеві продукти Ф. – фосфогліцеріновую кислоту і фосфогліцеріновий альдегід. Деякі трав'янисті, головним чином тропічного походження, рослини (наприклад, цукровий очерет, кукурудза, сорго) утворюють як перші продукти Ф. не трьох-, а чотирьохвуглецеві з'єднання – щавлевооцетову, яблучну і аспарагінову кислоти. Дорога автотрофної асиміляції Co 2 через фосфоєнолпіровіноградную кислоту, або фосфоєнолпіруват (ФЕП), з утворенням С 4 -дікарбонових кислот отримав назву С 4 -путі засвоєння вуглецю, а організми – С 4 -растеній. У листі таких рослин є два типи фотосинтезуючих кліток і Ф. йде в дві стадії. У клітках мезофіла аркуша відбувається первинна акцептація Co 2 на ФЕП за участю ФЕП-карбоксилазі, яка залучає Co 2 в реакції карбоксилювання навіть при дуже низьких концентраціях Co 2 в навколишньому повітрі. В результаті карбоксилювання утворюються щавлевооцетова, яблучна і аспарагінова кислоти. З них дві останні переходять в обкладочниє клітки провідних пучків аркуша, піддаються там декарбоксилюванню і створюють усередині кліток високу концентрацію Co 2 , що засвоюється вже через РДФ-карбоксилазу в циклі Калвіна. Це вигідно, по-перше, тому, що полегшує введення Co 2 в цикл Калвіна через карбоксилювання РДФ за допомогою РДФ-карбоксилазі, яка менш активна і вимагає для оптимальної роботи вищих концентрацій Co 2 , чим ФЕП-карбоксилаза. Крім того, висока концентрація Co 2 в обкладочних клітках зменшує світлове дихання (фотодихання ) і пов'язані з ним втрати енергії. Т. о. відбувається високоінтенсивний «кооперативний» Ф., вільний від зайвих втрат в світловому диханні, від кисневого інгібірування і добре пристосований до здійснення Ф. у атмосфері, бідною Co 2 і багатою O 2 .
Існують і ін. дороги перетворення Co 2 при Ф., в результаті яких в клітці в різних співвідношеннях утворюються різні органічні кислоти, білки і т.п. Співвідношення між цими групами з'єднань в рослині залежать від інтенсивності і якості світла, вигляду рослини і умов його розвитку (кореневого живлення, умов освітлення і ін.). Регулюючи умови розвитку рослин, можна управляти складом продуктів Ф. і тим самим – хімізмом рослини в цілому.
Роль фотосинтезу в біосфері. Поряд з Ф. на Землі здійснюються приблизно рівноцінні по масштабах, але протилежні по напряму процеси окислення органічних речовин і відновленого вуглецю при горінні паливних матеріалів (кам'яне вугілля, нафта, газ, торф, дрова і т.п.), при витрачанні органічних речовин живими організмами в процесі їх життєдіяльності (дихання, бродіння), в результаті яких утворюються повністю окислені з'єднання, – вуглекислий газ і вода, і звільняється енергія. Потім за допомогою енергії сонячної радіації вуглекислий газ, вода знову залучаються до процесів Ф. Т. о., енергія сонячного світла, використовувана при Ф., служить рушійною силою колосального по розмірах круговороту на Землі таких елементів, як вуглець, водень, кисень. У цей круговорот включаються і багато ін. елементи: N, S, Р, Mg, Ca і ін. За час існування Землі завдяки Ф. найважливіші елементи і речовини прошлі вже багато тисяч циклів повного круговороту.
В попередні епохи умови для Ф. на Землі були сприятливіші у зв'язку з сильною перевагою відновних процесів над окислювальними. Поступово величезні кількості відновленого вуглецю в органічних залишках виявилися похованими в надрах Землі, утворивши величезні поклади пальних копалин. В результаті цього в атмосфері сильно знизився відносний вміст вуглекислого газу (до 0,03 об'ємних %) і підвищився вміст кисню, що істотно погіршило умови для Ф.
Наслідком появи на Землі світу фотосинтезуючих рослин і безперервного новоутворення ними великих кількостей багатих енергією органічних речовин з'явився розвиток світу гетеротрофних організмів (бактерій, грибів, тварин, людини) – споживачів цих речовин і енергії. В результаті (в процесі дихання, бродіння, гниття, спалювання) органічні сполуки стали окислюватися і піддаватися розкладанню в таких же кількостях, в яких утворюють їх вищі рослини, водорості, бактерії. На Землі встановився круговорот речовин, в якому сума життя на нашій планеті визначається масштабами Ф. У поточному геологічному періоді (антропогеновом) розміри фотосинтетичної продуктивності на Землі, ймовірно, стабілізувалися. Проте у зв'язку з бурхливо зростаючим використанням продуктів Ф. основним її споживачем – людиною – доводиться думати про майбутнє виснаження пальних копалин, харчових, лісових ресурсів і т.п. Недостатня фотосинтетична потужність сучасної рослинності для регенерації атмосфери: рослинність Землі не здатна повністю засвоювати весь вуглекислий газ (відносний вміст його в атмосфері за останніх 100 років повільно, але неухильно зростає), що додатково поступає в довкілля в результаті бурхливий зростаючих масштабів видобутку і спалювання пальних копалин.
При цьому потенційна фотосинтетична активність рослин використовується далеко не повно. Проблема збереження, множення і найкращого використання фотосинтетичної продуктивності рослин – одна з найважливіших в сучасному природознавстві і практичній діяльності людини.
Фотосинтез і урожай. Одін з доріг підвищення загальної продуктивності рослин – посилення їх фотосинтетичної діяльності. Наприклад, щоб сформувати урожай пшениці в 40 ц/га, що складає 100 т загальної сухої біомаси, рослини повинні засвоїти близько 20 т Co 2 , фотохімічно розкласти близько 7,3 т H 2 O, виділити в зовнішнє середовище близько 13 т O 2 . Зазвичай за час вегетації рослин в середніх широтах (близько 3–4 мес ) на поверхню Землі приходить біля 2×10 9 ккал фотосинтетичний активній радіації (ФАР; в області спектру від 380 до 720 нм ) . З них в урожаї біомаси в 10 т запасається біля 40×10 6 ккал, тобто 2% ФАР. Остання енергія частково відбивається, але в більшій частині перетворюється на тепло і викликає випар величезних кількостей H 2 O. Т. о., для посилення фотосинтетичної діяльності рослин необхідно підвищити коефіцієнт використання рослинами сонячної радіації. Це досягається збільшенням в посівах розмірів листової поверхні, подовженням термінів активної діяльності листя, регулюванням густини стояння рослин. Важливе значення має спосіб розміщення рослин на площі (правильні норми висіву насіння), забезпечення їх достатньою кількістю Co 2 в повітрі, води, елементів грунтового живлення і т.д. Функціональна активність фотосинтетичного апарату, окрім зовнішніх умов, визначається також анатомічною будовою аркуша, активністю ферментних систем і типом метаболізму вуглецю. Велика роль належить селекції рослин – створенню сортів, що володіють високою інтенсивністю асиміляції Co 2 , і управлінню процесами, пов'язаними з ефективним використанням створюваних при Ф. органічних речовин. Важлива властивість високопродуктивних сортів – здатність використовувати велику частину асимілятів на формування коштовних в господарському відношенні органів (зерна в злаків, бульб в картоплі, коріння в коренеплодів і т.д.). З'ясування законів і основ фотосинтетичної продуктивності рослин, розробка принципів її оптимізації і підвищення – важливе завдання сучасності.
Літ.: Любіменко Ст Н,, Фотосинтез і хемосинтез на рослинному світі, М. – Л., 1935; Тімірязев До. А., Сонце, життя і хлорофіл, М., 1937 (Соч., т. 1–2); Годнев Т. Н., Будова хлорофілу і можливі дороги його освіти в рослині, М. – Л., 1947 (Тімірязевськоє читання. 7); Теренін А. Н., Фотохімія хлорофілу і фотосинтез, М., 1951 (Бахівське читання. 6); Рабіновіч Е., Фотосинтез, пер.(переведення) з англ.(англійський), т. 1–3 М., 1951–59; Нічипоровіч А. А., Фотосинтез і теорія здобуття високих урожаїв, М., 1956 (Тімірязевськоє читання, 15); Воськресенськая Н. П., Фотосинтез і спектральний склад світла, М., 1965; Андрєєва Т. Ф., Фотосинтез і азотний обмін листя, М., 1969; Теоретичні основи фотосинтетичної продуктивності, Сб. докл. на Междунар. симпозіумі, М., 1972; Сучасні проблеми фотосинтезу. До 200-ліття відкриття фотосинтезу, М., 1973; Красновський А. А., Перетворення енергії світла при фотосинтезі. Молекулярні механізми, М., 1974 (Бахівське читання. 29); Фотохімічні системи хлоропластів, До., 1975; Bioenergetics of photosynthesis, N. Y. – L. – Los Ang., 1975.