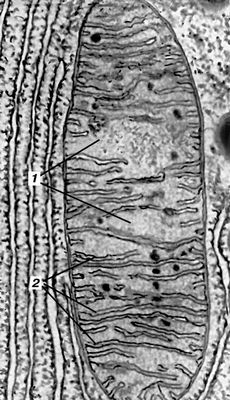
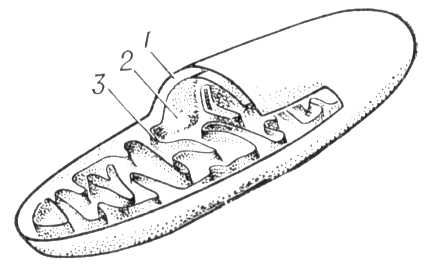
Мітохондрії (від греч.(грецький) mítos — нитка і chondríon — зернятко, крупинка), хондріосоми, постійно присутній в клітках тварин і рослин органоїд, що забезпечує клітинне дихання, в результаті якого енергія вивільняється або акумулюється в легко використовуваній формі (див. Окислення біологічне, Окислювальне фосфорилування ) . М. відсутні лише у бактерій, синезелених водоростей і інших прокаріотов, де їх функцію виконує клітинна мембрана. М. зазвичай концентруються у функціонально активних зонах клітки. Це округлі, палочковидні, гантелеобразниє і ін. освіти розміром зазвичай 0,5—1,5 мкм. Форма, число, розміри і функціональний стан М. міняються залежно від зовнішніх дій і фізіологічного стану клітки, а також при різних патологічних процесах. Число М. в клітках різних типів різне. Так, в клітці печінки щура їх близько 2500; у клітках з високою функціональною активністю (наприклад, в мотонейронах спинного мозку, в скелетному м'язі) число М. особливо велике. Біологічний напівперіод (оновлення половини складу) М. в клітках печінки складає 9,6—10,2 діб, в клітках бруньки — 12,4 діб. Спостережуване зазвичай впродовж життя клітки збільшення числа М. відбувається, мабуть, в результаті їх ділення. Внутрішня будова М. одноманітна. Дослідження за допомогою електронного мікроскопа показують, що основна речовина, або матрикс, М. оточено подвійною мембраною: зовнішньою — гладкою, і внутрішньою, — створюючою впячиванія, називається крістамі ( мал. 1 , 2 ). У мембранах М., що складаються із складних комплексів білків і ліпідів і організованих у вигляді дискретних субодиниць, сконцентрована більшість ферментів, що каталізують процеси окислення і фосфорилування. Зовнішня і внутрішня мембрани розрізняються великим або меншим вмістом деяких ліпідів і набором ферментів. Так, у внутрішній мембрані локалізовані ферментні комплекси, відповідальні за перенесення електронів і зв'язане з ним окислювальне фосфорилування; у зовнішній — фермент, що окислює моноаміни, і деякі ферменти обміну жирних кислот; у матриксі сконцентровані ферменти трікарбонових кислот циклу, окислення жирних кислот і ін., а в просторі між зовнішньою і внутрішньою мембранами — фермент аденілаткиназа, що каталізує оборотне перенесення фосфату з аденозинтрифосфорної кислоти (АТФ) на аденозінмонофосфорную кислоту (АМФ). Невід'ємні компоненти М. — дезоксирибонуклеїнова кислота (ДНК), розташована в матриксі у вигляді ниток і здібна до незалежної від ДНК(дезоксирибонуклеїнова кислота) ядра реплікації, і всі типи рибонуклеїнової кислоти (РНК). Крім того, в М. виявлені специфічні рибосоми (звані мітохондріальними), а також фермент, що каталізує ДНК(дезоксирибонуклеїнова кислота) -завісимий синтез РНК(рибонуклеїнова кислота). Присутність в М. всіх елементів системи біосинтезу білка забезпечує автономний синтез принаймні частини білків М. По сучасним виставам, М. є як би ендосимбіонтамі (див. Симбіогенез ) клітки, що має оформлене ядро. В процесі еволюції М. могли виникнути в результаті інфікування примітивних кліток аеробними бактерієподобнимі організмами, які потім пристосувалися до внутріклітинного існування і узяли на себе функцію дихання.
Літ.: Ленінджер А., Мітохондрія, пер.(переведення) з англ.(англійський) М., 1966; Алов І. А., Брауде А. І., Аспіз М. Е., Основи функціональної морфології клітки, 2 видавництва, М., 1969; Рудін Л., Уїлки Д., Біогенез мітохондрій, пер.(переведення) з англ.(англійський), М., 1970.