Дихання, сукупність процесів, які забезпечують потрапляння в організм кисню і виділення з нього вуглекислого газу (зовнішнє Д.) і використання кисню клітками і тканинами для окислення органічних речовин із звільненням енергії, що міститься в них, необхідної для життєдіяльності (тканинне дихання, клітинне Д.). Безкиснева дорога звільнення енергії властива лише невеликій групі організмів — так званим анаеробам (див. Бродіння ); в ході еволюції звільнення енергії в результаті Д. стало у переважної більшості організмів головним процесом, а анаеробні реакції збереглися в основному як проміжні етапи обміну речовин .
Д. тварин і людини. В простих, губок, кишечнополостних і деяких ін. організмів О 2 дифундує безпосередньо через поверхню тіла. З ускладненням організації і збільшенням розмірів тіла з'являються спеціальні дихання органи, а також система кровообігу, в якій циркулює рідина — кров або гемолімфа, що містить речовини, здатні зв'язувати і переносити O 2 і Co 2 (див. Гемоглобін ). У комах O 2 поступає в тканині з системи воздухоносних трубочок — трахей . У водних тварин, що використовують розчинений у воді O 2 , органами Д. служать зябра, забезпечені багатою мережею кровоносних судин. В цьому випадку O 2 , розчинений у воді, дифундує в кров, циркулюючу в судинах зябрових щілин. У багатьох риб значну роль грає кишкове Д., при якому повітря заковтується і O 2 поступає в кров через кровоносні судини кишечника; деяку роль в Д. риб грає також плавальний міхур; в тих, що багатьох мешкають у воді тварин обмін газів (головним чином СО 2 ) відбувається і через шкіру. В сухопутних тварин зовнішнє Д. забезпечується переважно легенями . В земноводних і багатьох ін. тварин поряд з цим функціонує шкірне Д. У птиць істотне значення мають ті, що сполучаються з легенями повітряні мішки, які змінюються в об'ємі при літальних рухах і полегшують Д. у польоті. В земноводних і плазуючих повітря в легені нагнітається рухами м'язів дна рота. У птиць, ссавців і людини зовнішнє Д. забезпечується ритмічною роботою дихальних м'язів (головним чином діафрагми і міжреберних м'язів), координованою нервовою системою. При скороченні цих м'язів об'єм грудної клітки збільшується і відбувається розтягування тих, що знаходяться в ній легенів; тому виникає різниця між атмосферним і внутрішньолегеневим тиском і повітря поступає в легені (вдих). Видих може бути пасивним — за рахунок спадання розтягнутої під час вдиху грудної клітки, а услід за нею і легенів; активний видих обумовлений скороченням деяких груп м'язів. Кількість повітря, що поступає в легені за 1 вдих, називається дихальним об'ємом (див. Легеневі об'єми ).
При Д. дихальна мускулатура долає еластичний опір, пов'язаний з пружністю грудної клітки, тягою легенів і поверхневим натягненням альвеол. Останнє, проте, значно знижується під впливом поверхнево активної речовини, альвеолярного епітелію, що виробляється клітками; тому альвеоли при видиху не спадаются, а при вдиху легко розширюються. Чим вище еластичний опір, тим важче розтягуються грудна клітка і легені; при глибокому Д. робота дихальної мускулатури, що витрачається на його подолання, різко зростає. Нееластичний опір Д. обумовлено головним чином тертям повітря при його русі по носових ходах, гортані, трахеї і бронхах. Воно залежить від швидкості потоку повітря під час Д. і від його характеру. При спокійному Д. потік близький до ламінарному (лінійному) в прямих ділянках воздухоносних доріг і до турбулентного (вихровому) в місцях розгалуження або звуження. Із збільшенням швидкості потоку (при форсованому Д.) турбулентність зростає і для просування повітря потрібна вища різниця тиску, а отже, і збільшення роботи дихальних м'язів. Нерівномірний розподіл опору руху повітря по дихальних дорогах приводить до того, що вступ повітря в різні групи легеневих альвеол відбувається нерівномірно; така різниця у вентиляції особливо значительна при легеневих захворюваннях.
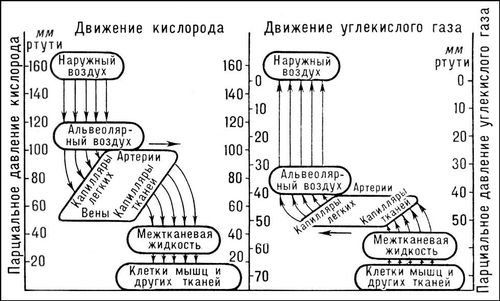
Кількість повітря, що вентилює легені в 1 мін , називається хвилинним об'ємом дихання (МОД). МОД дорівнює твору дихального об'єму на частоту Д. (число дихательних рухів в 1 мін , рівне у людини приблизно 15—18) і складає у дорослої людини у спокої 5—8 л/мін . Лише частина МОД (близько 70%) бере участь в обміні газів між вдихуваним і альвеолярним повітрям, цю частину називають об'ємом альвеолярної вентиляції; остання частина МОД використовується для «промивання» так званого мертвого, або шкідливого, простору дихальних доріг, в якому на початок видиху зберігається зовнішнє повітря, що заповнило його в кінці передуючого вдиху (об'єм мертвого простору близько 160 мл ). Вентиляція альвеол забезпечує постійність складу альвеолярного повітря . Парціальний тиск O 2 ( p O2 ) і Co 2 ( p Co2 ) в альвеолярному повітрі вагається в дуже вузьких межах і складає для О 2 близько 13 кн/м 2 (100 мм рт. ст. ) і для СО 2 близько 5,4 кн/м 2 (40 мм рт. ст. ).
Обмін газів між альвеолярним повітрям і венозною кров'ю, що поступає в капіляри легенів, здійснюється через альвеоло-капілярну мембрану, загальна поверхня якої дуже велика (у людини близько 90 м 2 ). Дифузія O 2 в кров забезпечується різницею парціального тиску O 2 в альвеолярному повітрі і у венозній крові (8—9 кн/м 2 , або 60—70 мм рт. ст. ). Co 2 , приношуваний кров'ю з тканин в зв'язаній формі (бікарбонати, солі вугільної кислоти і карбгемоглобін), звільняється в капілярах легенів за участю ферменту карбоангидрази і дифундує з крові в альвеоли; різниця p Co2 між венозною кров'ю і альвеолярним повітрям складає близько 7 мм рт. ст. Здатність альвеолярної стінки пропускати O 2 і Co 2 , так звана дифузійна здатність легенів, дуже велика: у спокої вона складає в 1 мін приблизно 30 мл O 2 на 1 мм різниці p O2 між альвеолярним повітрям і кров'ю; для Co 2 ця величина у багато разів більше. Тому парціальний тиск газів в артеріальній крові, що відтікала з легенів, встигає наблизитися до їх тиску в альвеолярному повітрі. Перехід O 2 в тканини і видалення з них Co 2 також відбуваються шляхом дифузії, т.к. p O2 в тканинній рідині 2,7—5,4 кн/м 2 (20—40 мм рт. ст. ), а в клітках ще нижче, а p Co2 в клітках може досягати 60 мм рт. ст. (см. мал.(малюнок) ).
Вжиток O 2 клітками і тканинами і освіта ними Co 2 , що складає суть тканинного, або клітинного, Д., — одна з основних форм дисиміляція, що здійснюється у тварин і рослин в принципі однаково. Високий вжиток O 2 характерний для тканин нирок, кори великих півкуль головного мозку, серця. У результатеокислювально-відновних реакцій тканинного Д. звільняється енергія, що витрачається на всі життєві прояви. Процес цей здійснюється в мітохондріях і складається з дегідрування субстратів Д. — вуглеводів і продуктів їх розщеплювання, жирів і жирних кислот, амінокислот і продуктів їх дезамінування. Субстрати Д. поглинають O 2 і служать джерелом Co 2 (відношення —
називається дихальним коефіцієнтом ). Енергія, що звільняється при окисленні органічних речовин, не використовується тканинами безпосередньо, т.к. около 70% її витрачається на освіту АТФ — одній з аденозінфосфорних кислот подальше ферментативне розщеплювання якої забезпечує енергетичні потреби тканин, органів і організму в цілому (див. Окислення біологічне, Окислювальне фосфорилування ). Т. о., з біохімічної точки зору Д. — це перетворення енергії вуглеводів і ін. речовин на енергію макроергічних фосфатних зв'язків.
Постійність p O2 і p Co2 в альвеолярному повітрі, а отже, і в артеріальній крові може підтримуватися лише за умови, якщо альвеолярна вентиляція відповідає швидкості вжитку організмом O 2 і утворення Co 2 , тобто рівню обміну речовин. Ця умова забезпечується завдяки досконалим механізмам регуляції Д. Управленіє частотою і глибиною Д. здійснюється рефлекторним дорогою. Так, підвищення p Co2 і зниження p O2 в альвеолярному повітрі і в артеріальній крові збуджують хеморецептори синокаротідной і кардіоаортальной зон, що приводить до збудження дихального центру і збільшенню МОД. Згідно з класичними виставами, підвищення p Co2 в артеріальній крові, що омиває дихальний центр також збуджує його і викликає збільшення МОД. Т. о., регуляція Д. по відхиленню p O2 і p Co2 в артеріальній крові, здійснювана за типом зворотного зв'язку, забезпечує оптимальний МОД. Проте у ряді випадків, наприклад при м'язовій роботі, МОД збільшується до настання в обміні речовин зрушень, які приводять до змін в газовому складі крові. Це посилення вентиляції обумовлене сигналами, що поступають в дихальний центр від рецепторів рухового апарату, рухової зони кори великих півкуль мозку, а також умовними рефлексами на різні сигнали, пов'язані із звичною роботою і її обстановкою. Т. о., управління Д. здійснюється складною самонавчальною системою не лише за принципом регулювання по відхиленню, але і по сигналах, застережливих про можливі відхилення. Зміна вдиху і видиху забезпечується системою взаимодоповнюючих механізмів. Під час вдиху в дихальний центр по волокнах блукаючих нервів поступають імпульси від рецепторів розтягування, що знаходяться в легенях. При досягненні легенями певного об'єму ця імпульсация гальмує клітки дихального центру, збудження яких викликає вдих. При виключенні нервових доріг, що забезпечують вступ імпульсів в дихальний центр, ритмічність Д. зберігається завдяки автоматизму центру, проте характер ритму різко відрізняється від нормального. При порушеннях Д. і механізмів його регуляції виникають зміни газового складу крові (див. Гіпоксія ).
Методи дослідження Д. всілякі. У фізіології праці і спорту, клінічній медицині широко застосовують реєстрацію глибини і частоти дихальних рухів, виміри газового складу повітря, що видихається, артеріальної крові, плеврального і альвеолярного тиску. Див. також Газообмін .
Літ.: Сеченов І. М., Ізбр. праці, М., 1935; Холден Дж. і Прістлі Дж., Дихання, пер.(переведення) з англ.(англійський), М-кодом.—Л., 1937; Маршак М. Е., Регуляція дихання у людини, М., 1961; Фізіологія людини, М., 1966; Comroe J. Н., Physiology of respiration, Chi., 1966; Dejours P., Respiration, Oxf., 1966.
Л. Л. Шик.
Д. рослин. Д. властиво всім органам, тканинам і кліткам рослини. Про інтенсивність Д. можна судити, вимірюючи кількість що виділяється тканиною Co 2 або що поглинається нею O 2 . Інтенсивніше дихають молоді, швидко зростаючі органи і тканини рослин. Найактивніше Д. репродуктивних органів, потім листя; слабкіше Д. стебел і коріння. Тіньовитривалі рослини дихають слабкіше світлолюбних. Для високогірних рослин, адаптованих до зниженого парціального тиску O 2 , характерна підвищена інтенсивність Д. Очень активно Д. плісневих грибів, бактерій. Д. посилюється з підвищенням температури (на кожні 10°С — приблизно в 2—3 рази), припиняючись при 45—50°С. У тканинах зимуючих органів рослин (нирки листяних дерев, голки хвойних) Д. продовжується (з різко пониженою інтенсивністю) і при значних морозах. Д. стимулюють механічні і хімічні роздратування (пораненія, деякі отрути, наркотики і т.п.). Закономірно змінюється Д. в ході розвитку рослини і його органів. Сухе (що покояться) насіння дихає дуже слабо; при набуханні і подальшому проростанні насіння Д. посилюється в сотні і тисячі разів. Із закінченням періоду активного зростання рослин Д. їх тканин слабшає, що пов'язане з процесом старіння протоплазми. При дозріванні насіння, плодів інтенсивність Д. зменшується.
Згідно теорії радянського біохіміка А. Н. Баха, процес Д., тобто окислення вуглеводів, жирів, білків, здійснюється за допомогою окислювальної системи клітки в два етапи: 1) активування O 2 повітря шляхом його приєднання до тих, що містяться в живій клітині ненасиченим, здатним мимоволі окислюватися з'єднанням (оксигеназам) з утворенням перекисів; 2) активування останніх із звільненням атомарного кисню, здатного окислювати важко окислювані органічні речовини. По теорії дегідрування російського ботаніка Ст І. Палладіна, найважливіша ланка Д. — активація водню субстрата, здійснювана дегідрогеназою . Обов'язковий учасник складного ланцюга процесів Д. — вода, водень якої разом з воднем субстрата використовується для відновлення з'єднань, що самоокисляющихся, — так званих дихальних пігментів. Co 2 , що виділяється при Д., утворюється без участі кисню повітря, тобто анаеробний. Кисень повітря йде на окислення дихальних хромогенів, що перетворюються при цьому на дихальні пігменти. Подальший розвиток теорія Д. отримала в дослідженнях радянського ботаніка С. П. Костичева, згідно з якими перші етапи аеробного Д. аналогічні процесам, властивим анаеробам. Перетворення проміжного продукту, що утворюється при цьому, можуть йти за участю кисню, що властиве аеробам. В анаеробів же ці перетворення йдуть без участі молекулярного кисню. По сучасних виставах, процес окислення, який складає хімічну основу Д., полягає у втраті речовиною електрона. Здатність приєднувати або віддавати електрони залежить від величини окислювального потенціалу з'єднання. Кисень володіє найвищим окислювальним потенціалом і, отже, максимальною здатністю приєднувати електрони. Проте потенціал кисню сильно відрізняється від потенціалу дихального субстрата. Тому роль проміжних переносників електронів від дихального субстрата до кисню виконують специфічні з'єднання. Поперемінно окислюючись і відновлюючись, вони утворюють систему перенесення електронів. Приєднавши до себе електрон від менш окисленого компонента, такий переносник відновлюється і, віддаючи його наступному компоненту з вищим потенціалом, окислюється. Так електрон передається від однієї ланки дихального ланцюга до іншого і, врешті-решт, кисню. Такий завершальний етап Д.
Всі ці процеси (активація кисню, водню, перенесення електрона по ланцюгу на кисень) здійснюються головним чином в мітохондріях завдяки розгалуженій системі окислювально-відновних ферментів (див. Цитохроми ). По шляху дотримання до кисню електрони, мобілізуємиє спочатку від молекули органічної речовини, поступово віддають укладену в них енергію, яку клітка запасає у формі хімічних сполук, головним чином АТФ.
Завдяки досконалим механізмам запасання і використання енергії процеси енергообміну в клітці йдуть з дуже високим ккд(коефіцієнт корисної дії), поки недосяжним в техніці. Біологічна роль Д. не вичерпується використанням енергії, увязненою в окислюваній органічній молекулі. В ході окислювальних перетворень органічних речовин утворюються активні проміжні з'єднання — метаболіти, які жива клітина використовує для синтезу специфічних складових частин своєї протоплазми, утворення ферментів і ін. Всім цим визначається центральне місце, займане Д. у комплексі процесів обміну речовин живої клітини. У Д. схрещуються і ув'язуються процеси обміну білків, нуклеїнових кислот, вуглеводів, жирів і ін. компонентів протоплазми.
Літ.: Костичев С. П., Фізіологія рослин, 3 видавництва, т. 1, М-код.—Л., 1937; Бах А. Н., Собр. праць по хімії і біохімії, М., 1950; Таусон Ст О., Основні положення рослинної біоенергетики, М-код.—Л., 1950; Джеймс Ст О., Дихання рослин, перло.(переведення) з англ.(англійський), М., 1956; Палладін Ст І., Ізбр. праці, М., 1960; Міхлін Д. М., Біохімія клітинного дихання, М., 1960; Сент-Дьердьі А., Біоенергетика, пер.(переведення) з англ.(англійський), М., 1960; Рубін Би. А., Ладигина М. Е., Ензімология і біологія дихання рослин, М., 1966; Рекер Е., Біоенергетичні механізми, пер.(переведення) з англ.(англійський), М., 1967; Рубін Би. А., Курс фізіології рослин, 3 видавництва, М., 1971; Кретовіч Ст Л., Основи біохімії рослин, М., 1971.