Газообмін (біологічне), обмін газів між організмом і зовнішнім середовищем. З довкілля в організм безперервно потрапляє кисень, який споживається всіма клітками, органами і тканинами; з організму виділяються вуглекислий газ, що утворюється в нім, і незначна кількість ін. газоподібних продуктів обміну речовин . Р. необхідний майже для всіх організмів, без нього неможливий нормальний обмін речовин і енергії, а отже і саме життя.
Кисень, що поступає в тканині, використовується для окислення продуктів, що утворюються у результаті довгому ланцюгу хімічних перетворень вуглеводів, жирів і білків. При цьому утворюються СО 2 , вода, азотисті з'єднання і звільняється енергія, використовувана для підтримки температури тіла і виконання роботи. Кількість що утворюється в організмі і зрештою виділяється з його СО 2 залежить не лише від кількості споживаного О 2 , але і від того, що переважно окислюється: вуглеводи, жири або білки. Відношення що видаляється з організму СО 2 до поглиненого за той же час О 2 називається дихальним коефіцієнтом, який дорівнює приблизно 0,7 при окисленні жирів, 0,8 при окисленні білків і 1,0 при окисленні вуглеводів. Кількість енергії, що звільняється на 1 л спожитого О 2 (калорічеський еквівалент кисню), рівне 20,9 кдж (5 ккал ) при окисленні вуглеводів і 19,7 кдж (4,7 ккал ) при окисленні жирів. Т. о., по вжитку О 2 в одиницю часу і по дихальному коефіцієнту можна розрахувати кількість енергії, що звільнилася в організмі.
Р. (відповідно і витрата енергії) в пойкілотермних тварин (холоднокровних) знижується з пониженням температури тіла. Така ж залежність виявлена і в гомойотермних тварин (теплокровних) при виключенні терморегуляції (в умовах природної або штучної гіпотермії ) ; при підвищенні температури тіла (при перегріві, різних захворюваннях) Р. збільшується.
При пониженні температури довкілля Р. у теплокровних тварин (особливо в дрібних) збільшується в результаті збільшення теплопродукциі . Р. збільшується також після їди, особливо багатої білками (т.з. специфічно-динамічна дія їжі). Найбільших величин Р. досягає при м'язовій діяльності. У людини при роботі помірної потужності Р. збільшується, через 3—6 мін після її початку досягає певного рівня і потім стримується протягом всього часу роботи на цьому рівні. При роботі великої потужності Р. безперервно зростає; незабаром після досягнення максимального для даної людини рівня (максимальна аеробна робота) роботу доводиться припиняти, т. до. потребность організму в О 2 перевищує цей рівень. Спочатку після закінчення роботи зберігається підвищений вжиток О 2 використовуваного для покриття кисневого боргу, тобто для окислення продуктів обміну речовин, що утворилися під час роботи. Вжиток О 2 може збільшуватися з 200—300 мл/мін в стані спокою до 2000—3000 при роботі, а у добре тренованих спортсменів — до 5000 мл/мін. Відповідно збільшуються виділення СО 2 і витрата енергії; одночасно відбуваються зрушення дихального коефіцієнта, пов'язані із змінами обміну речовин, кислотно-лужної рівноваги і легеневої вентиляції.
Розрахунок загальної добової витрати енергії у людей різних професій і способу життя, заснований на визначеннях Р., важливий для нормування живлення. Дослідження змін Р. при стандартній фізичній роботі застосовуються у фізіології праці і спорту, в клініці для оцінки функціонального стану систем, що беруть участь в Р.
Порівняльна постійність Р. при значних змінах парціального тиску О 2 в довкіллі, порушеннях роботи органів дихання і т. п. забезпечується пристосовними (компенсаторними) реакціями систем, що беруть участь в Р. і регульованих нервовою системою.
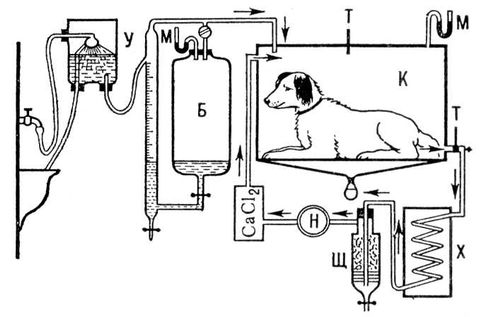
Р. у людини і тварин прийнято досліджувати в умовах повного спокою, натщесерце, при комфортній температурі середовища (18—22 °С). Кількості споживаного при цьому О 2 і що звільняється енергії характеризують основний обмін . Для дослідження Р. застосовуються методи, засновані на принципі відкритої або закритої системи. У першому випадку визначають кількість повітря, що видихається, і його склад (за допомогою хімічних або фізичних газоаналізаторів), що дозволяє обчислювати кількості споживаного О 2 і СО 2 , що виділяється . У другому випадку дихання відбувається в закритій системі (герметичній камері або із спірографа, сполученого з дихальними дорогами), в якій поглинається СО, що виділяється, 2 , а кількість спожитого з системи О 2 визначають або виміром рівної йому кількості що автоматично поступає в систему О 2 , або по зменшенню об'єму системи ( мал. ).
Літ.: Гинецинський А. Р., Лебединський А. Ст, Курс нормальної фізіології, М. 1956; Фізіологія людини, М., 1966, с. 134—56; Берковіч Е. М., Енергетичний обмін в нормі і патології, М., 1964 (є бібл.); Проссер Л., Браун Ф., Порівняльна фізіологія тварин, пер.(переведення) з англ.(англійський), М., 1967, с. 186—237.