Гемоглобін (Hb) (від гемо... і лат.(латинський) globus — куля), червоний залізовмісний пігмент крові людини, хребетних і деяких безхребетних тварин; у організмі виконує функцію перенесення кисню (O 2 ) з органів дихання до тканин; грає також важливу роль в перенесенні вуглекислого газу від тканин в органи дихання. У більшості безхребетних Р. вільно розчинений в крові; в хребетних і деяких безхребетних знаходиться в червоних кров'яних клітках — еритроцитах, складаючи до 94% їх сухого залишку. Молярна маса Р., включеного в еритроцити, близько 66 000, розчиненого в плазмі — до 3000000. По хімічній природі Р. — складний білок — хромопротєїд, що складається з білка глобіну і железопорфіріна — гема. У вищих тварин і людини Р. складається з 4 субодиниць-мономерів з молярною масою близько 17000; два мономери містять по 141 залишку амінокислот ( а -цепі), два інших — по 146 залишків ( b -цепі).
Просторові структури цих поліпептидів багато в чому аналогічні. Вони утворюють характерні «гідрофобні кишені», в яких розміщені молекули гема (по одній на кожну субодиницю). З 6 координаційних зв'язків атома заліза, що входить до складу гема, 4 направлені на азот піррольних кілець; 5-я сполучена з азотом імідазольного кільця гістидину, що належить поліпептидам і що стоїть на 87-м-коді місці в а -цепі і на 92-м-коді місці в b -цепі; 6-я зв'язок направлений на молекулу води або ін. групи (ліганди) і у тому числі на кисень. Субодиниці рихло зв'язані між собою водневими, сольовими і ін. нековалентними зв'язками і легко диссоціюють під впливом амідов, підвищеній концентрації солей з освітою головним чином симетричних димірів ( ab ) і частково a- і b -мономеров. Просторова структура молекули Р. вивчена методом рентгеноструктурного аналізу (М. Перуц, 1959).
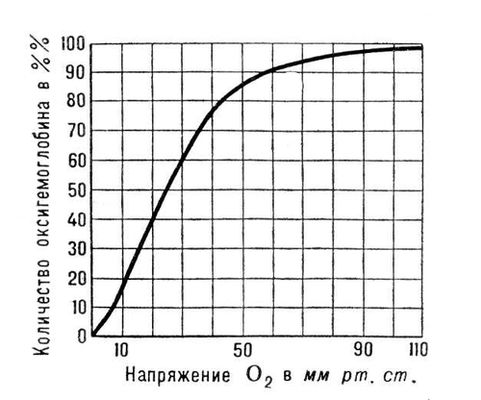
Послідовність розташування амінокислот в a- і b -цепях Р. ряду вищих тварин і людини повністю з'ясована. У зібраній в тетрамер молекулі Р. все 4 залишки гема розташовано на поверхні і легко доступні реакції з O 2 . Приєднання O 2 забезпечується вмістом в гемі атома Fe 2+ . Ця реакція обратіма і залежить від парціального тиску (напруга) O 2 . У капілярах легенів, де напруга O 2 близько 100 мм рт . ст. , Р. з'єднується з O 2 (процес оксигенації), перетворюючись на оксигенірованний Р. — оксигемоглобін. У капілярах тканин, де напруга O 2 значно нижче (ок. 40 мм рт . ст. ), відбувається дисоціація оксигемоглобіну на Р. і O 2 ; останній поступає в клітки органів і тканин, де парціальний тиск O 2 ще нижче (5—20 мм рт . cm. ); в глибині кліток воно падає практично до нуля. Приєднання O 2 до Р. і дисоціація оксигемоглобіну на Р. і O 2 супроводяться конформаційними (просторовими) змінами молекули Р., а також його оборотним розпадом на диміри і мономери з подальшою агрегацією в тетрамери.
Змінюються при реакції з O 2 і ін. властивості Г.: оксигенірованний Р. — в 70 разів сильніша кислота, ніж Г. Ето грає велику роль в скріпленні в тканинах і віддачі в легенях Co 2 . Характерні смуги поглинання у видимій частині спектру: в Р. — один максимум (при 554 ммк ), в оксигенірованного Р. — два максимуми при 578 і 540 ммк . Р. здатний безпосередньо приєднувати Co 2 (у результаті реакції Co 2 з Nh 2 -rpyппамі глобіну); при цьому утворюється карбгемоглобін — з'єднання нестійке, таке, що легко розпадається в капілярах легенів на Р. і Co 2 .
Кількість Р. в крові людини — в середньому 13—16 г% (або 78%—96% по Салі); у жінок Р. декілька менше, ніж у чоловіків. Властивості Р. міняються в онтогенезі. Тому розрізняють Р. ембріональний, Р. — плоду (foetus) — HBF, Р. дорослих (adult) — HBA. Спорідненість до кисню в Р. плоду вище, ніж в Р. дорослих, що має істотне фізіологічне значення і забезпечує велику стійкість організму плоду до недоліку O 2 . Визначення кількості Р. в крові має важливе значення для характеристики дихальної функції крові в нормальних умовах і при самих різних захворюваннях, особливо при хворобах крові. Кількість Р. визначають спеціальними приладами — гемометрами.
При деяких захворюваннях, а також при природжених аномаліях крові (див. Гемоглобінопатії ) в еритроцитах з'являються аномальні (патологічні) Р., такі, що відрізняються від нормальних заміщенням амінокислотного залишку в ( - або b -цепях. Виділено більше 50 різновидів аномальних Р. Так, при серповидноклітинній анемії виявлений Р., в b -цепях якого глутамінова кислота, що стоїть на 6-м-коді місці від n-кінця, заміщена валіном. Аномалії еритроцитів, пов'язані з вмістом гемоглобіну F або Н, лежать в основі таласемії, метгемоглобінемії . Дихальна функція деяких аномальних Р. різко порушена, що обумовлює різні патологічні стани (анемії і ін.). Властивості Р. можуть мінятися при отруєнні організму, наприклад чадним газом, що викликає утворення карбоксигемоглобіну, або отрутами, переводячими Fe 2+ гема в Fe 3+ з утворенням метгемоглобіну. Ці похідні Р. не здатні переносити кисень. Р. різних тварин володіють видовою специфічністю, обумовленою своєрідністю будови білкової частини молекули. Р. що звільняється при руйнуванні еритроцитів, — джерело утворення жовчних пігментів .
В м'язовій тканині міститься м'язовий Р. — міоглобін, по молярній масі, складу і властивостям близький до субодиницям Р. (мономерам). Аналоги Р. виявлені в деяких рослин (наприклад, леггемоглобін міститься в бульбах бобів).
Літ.: Коржуєв П. А., Гемоглобін, М., 1964; Гауровіц Ф., Хімія і функції білків, пер.(переведення) з англ.(англійський), 2 видавництва, М., 1965, с. 303—23; Інгрем Ст, Біосинтез макромолекул, пер.(переведення) з англ.(англійський), М., 1966, с. 188—97; Рапопорт С. М., Медична біохімія, пер.(переведення) з йому.(німецький), М., 1966; Перутц М., Молекула гемоглобіну, в збірці: Молекули і клітки, М., 1966; Цукеркандль Е.; Еволюція гемоглобіну, там же; Fanelli A. R., ANTONINIE., Caputo A., Hemoglobin and myoglobin, «Advances in Protein Chemistry», 1964, v. 19, р. 73—222; Antonini Е., Brunori M., Hemoglobin, «Annual Review of Biochemistry», 1970, v. 39, р. 977—1042.