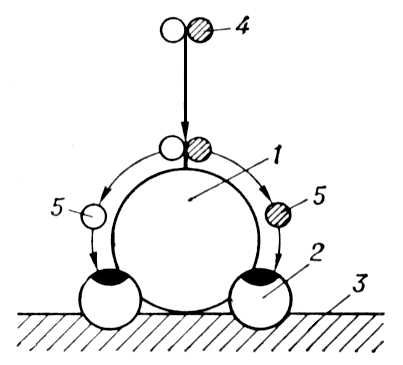
Травлення, сукупність процесів, що забезпечують механічне подрібнення і хімічне (головним чином ферментативне) розщеплювання харчових речовин на компоненти, позбавлені видовій специфічності і придатні до всмоктування і участі в обміні речовин організму тварин і людини. Що потрапляє в організм їжа всесторонньо обробляється під дією різних травних ферментів, що синтезуються спеціалізованими клітками, причому розщеплювання складних харчових речовин (білків, жирів і вуглеводів) на все дрібніші фрагменти відбувається з приєднанням до них молекули води (див. Гідроліз ). Білки розщеплюються зрештою на амінокислоти, жири — на гліцерин і жирні кислоти, вуглеводи — на моносахариди. Ці відносно прості речовини піддаються всмоктуванню, а з них в органах і тканинах знов синтезуються складні органічні сполуки. Відомо 3 основних типа П.: внутріклітинне, позаклітинне (дістантноє) і мембранне ( мал. 1 ).
Внутріклітинне П.: нерозщеплений або неповністю розщеплений харчовий субстрат поступає всередину клітки, де піддається подальшому гідролізу ферментами цитоплазми. Такий еволюційно більш древній тип П. поширений у всіх одноклітинних, в деяких нижчих багатоклітинних організмів (наприклад, в губок) і у вищих тварин. У останньому випадку маються на увазі фагоцитарні властивості білих кров'яних кліток (див. Лейкоцити ) і ретікуло-ендотеліальної системи, а також один з різновидів фагоцитозу — так званий піноцитоз, властивий кліткам екто- і ентодермального походження. Внутріклітинне П. може бути реалізоване не лише в цитоплазмі, але і в спеціальних внутріклітинних порожнинах — травних вакуолях, що існують постійно або що утворюються при фаго- і піноцитозі. Передбачається, що в внутріклітинному П. можуть брати участь лізосоми, ферменти яких поступають в травні вакуолі.
Позаклітинне, або дістантноє, П.: ферменти, що синтезуються в клітках, переносяться в позаклітинне середовище організму і здійснюють свою дію на відстані від секретуючих кліток. Позаклітинне П. переважає у кільчастих черв'яків, ракоподібних, комах, головоногих, покривників і хордових окрім ланцетника. У більшості високоорганізованих тварин секреторні клітки розташовані досить далеко від порожнин, де реалізується дія травних ферментів (слинні залози і підшлункова залоза у ссавців). Якщо дістантноє П. відбувається в спеціальних порожнинах, прийнято говорити про порожнинний П. П. Дістантноє може проходіть за межами організму, що продукує ферменти. Так, при дістантном внеполостном П. комахи вводять травні ферменти в обездвіженную видобуток, а бактерії виділяють всілякі ферменти в культуральне середовище.
Мембранне, або прістеночноє, П. здійснюється ферментами, локалізованими на структурах клітинної мембрани, і займає проміжне положення між позаклітинним і внутріклітинним. У більшості високоорганізованих тварин таке П. відбувається на поверхні мембран мікроворсинок кишкових кліток і є основним механізмом проміжних і завершальних стадій гідролізу. Мембранне П. забезпечує досконале сполучення травних і транспортних процесів і їх максимальне зближення у просторі та часі. Це досягається в результаті спеціальної організації травних і транспортних функцій клітинної мембрани у вигляді своєрідного травно-транспортного «конвеєра», сприяючого передачі кінцевих продуктів гідролізу з ферменту на переносник або вхід в транспортну систему ( мал. 2 ). Мембранне П. виявлене у людини, ссавців, птиць, земноводних, риб, круглоротих і багатьох представників безхребетних тварин (комахи, ракоподібні, молюски, черв'яки). Кожному з 3 типів П. властиві як певні переваги, так і обмеження. У процесі еволюції більшість організмів стали поєднувати ці процеси; частіше вони комбінуються в одного і того ж організму, що сприяє оптимальній ефективності і економічності травної системи .
У людини, вищих і багатьох нижчих тварин травний апарат підрозділяють на ряд відділів, що виконують специфічні функції: 1) що сприймає; 2) провідний який в деяких видів тварин розширений з утворенням спеціального депо; 3) травні відділи — а) роздрібнення їжі і початкових етапів П. (в деяких випадках воно завершується в цьому відділі), би) подальшого П. і всмоктування; 4) всмоктування води; цей відділ має особливе значення для наземних тварин, в нім всмоктується велика частина води, що поступає в кишечник (англійський учений Дж. Дженнінгс, 1972). У кожному з відділів харчова маса, залежно від її властивостей і спеціалізації відділів, затримується на певний час або переводиться в наступний відділ.
Травлення в ротовій порожнині. У ссавців, більшості ін. хребетних і багатьох безхребетних тварин їжа піддається в ротовій порожнині (у людини вона знаходиться тут в середньому 10—15 сік ) як механічному подрібненню дорогою жування, так і первинній хімічній обробці під дією слини, яка, змочуючи харчову масу, забезпечує формування харчової грудки. Хімічна обробка їжі в роті полягає в основному в переварюванні (у людини і всеїдних) вуглеводів амілазою слини. Тут же (головним чином на мові) розташовані смакові органи, що здійснюють дегустацію їжі. За допомогою рухів мови і щік харчова грудка подається на корінь мови і в результаті ковтання поступає в стравохід, а потім в шлунок.
Травлення в желудке . Їжа накопичується в шлунку, перемішується і просочується кислим шлунковим соком, що володіє ферментативною активністю, вираженими антибактеріальними властивостями і здатністю денатурувати клітинні структури. Основна функція шлунку: депонування їжі, її механічна і хімічна обробка, що включає початкові стадії П. (головним чином білків під дією протеолітичних ферментів ), а також поступова евакуація харчової маси в кишечник . В шлунку їжа знаходиться залежно від її кількості і складу від 4 до 10 до (у людини в середньому 3,5—4 ч ) . У багатьох тварин шлунок має декілька відділів, що виконують різні функції. Наприклад, в жуйних в шлунку відбуваються основні перетворення харчової маси під впливом діяльності бактерій і простих. Слизиста оболонка шлунку секретує неактивний пепсиноген, що активується у присутності соляної кислоти і трансформований в активний пепсин, що здійснює початкові стадії гідролізу білків, а також парапепсини, гастріксин, желатіназу (у природних умовах що розщеплює, мабуть, колаген сполучні тканини) і катепсини, що беруть участь в шлунковому П. на ранніх етапах онтогенетичного розвитку. У шлунковому соку деяких жуйних в період молочного живлення виявляється реннін, або хімозин, що викликає звурджування і подальше розщеплювання казеїну і що діє, на відміну від пепсину, в слабокислому або нейтральному середовищі. У шлунковому соку присутнє невелике кількість ліпази, роль якої, проте, невелика. Амілаза слини до її денатурації соляною кислотою продовжує розщеплювання вуглеводів, що почалося в порожнині рота. У порожнині шлунку діють також ферменти підшлункового соку, що закидається антиперистальтичними рухами, головним чином при прийомі жирної їжі.
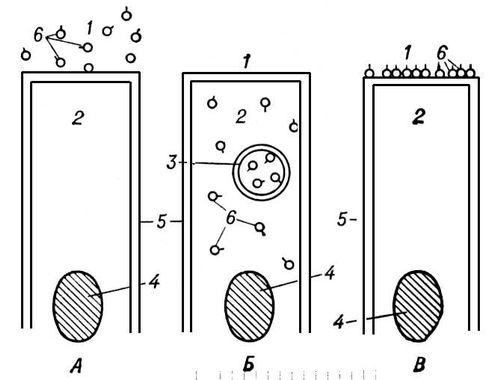
Травлення в кишечнике . З шлунку харчова маса порціями поступає в кишечник, де найінтенсивніше (особливо в початковій частині тонкої кишки) відбуваються процеси ферментативного гідролізу і перехід до всмоктування. Фаза П. в тонкому кишечнику реалізується в середовищі, близькому до нейтральної. Перехід від первинного переварювання в кислому середовищі (шлунок) до переварювання в нейтральній або слаболужній (тонка кишка) типовий як для людини і вищих тварин, так і для нижчих багатоклітинних і одноклітинних організмів, в яких в травних вакуолях підтримується спочатку кисла, а потім лужна реакція. Більшість надмолекулярних агрегацій і крупних молекул (білки і продукти їх неповного гідролізу, вуглеводи і жири) у людини і вищих тварин розщеплюються в порожнині тонкої кишки переважно під дією ферментів, що секретуються підшлунковою залозою і що поступають в дванадцятипалу кишку. Пептиди, що утворилися під дією пепсину шлунку, і нерозщеплені білки гидролізуются протеазами підшлункового соку: трипсином, хімотрипсином, карбоксипептидазою і еластазою. В результаті послідовної дії цих ферментів в порожнині тонкої кишки з крупних білкових молекул і поліпептидів утворюються низькомолекулярні пептиди і незначне кількість амінокислот. Вуглеводи (крохмаль і глікоген) гидролізуются під впливом а-амілази підшлункового соку, що розщеплює їх до трі- і дісахарідов без значного накопичення глюкози. У гідролізі жирів істотну роль грає жовч, що виділяється печиву . Жовч активує ліпазу підшлункового соку і емульгує жири, що приводить до збільшення поверхні зіткнення їх з ліпазою, розчиненою у водній фазі. У порожнині тонкої кишки цей фермент поетапно відщеплює жирні кислоти і приводить до утворення ді- і моногліцеридів і незначної кількості вільних жирних кислот і гліцерину. Продукти гідролізу, що утворюються, в результаті перемішуючих рухів кишкової мускулатури (див. Маятникоподібні рухи ) стикаються з поверхнею кишки, де відбувається подальша їх обробка шляхом мембранного П. ( мал. 3 ) . У зв'язку з вираженою поверхневою активністю продукти гідролізу поступають в зону щіткової облямівки (якщо розміри їх молекул не дуже великі), чому сприяє їх перенесення в потоках розчинника, що виникають в результаті всмоктування води кишковими клітками.
Проміжні і завершальні стадії П. реалізуються ферментами, локалізованими на поверхні мембран кишкових кліток, де починається всмоктування. У мембранному П. беруть участь: 1) ферменти підшлункового соку (а-амілаза, ліпаза, трипсин, хімотрипсин, еластаза і ін.), адсорбовані в різних шарах так званого глікокалікса, що покриває мікроворсинки і мукополісахарідную, що є, тривимірну мережу; 2) власне кишкові ферменти (g-амілаза, оліго- і дісахарідази, різні тетра-, трі- і дипептидаза, амінопептидаза, лужна фосфатаза і її ізоензіми, моногліцерідліпаза і ін.), синтезовані клітками кишкового епітелію і переносимі на поверхню їх мембран, де вони здійснюють травні функції. Адсорбовані ферменти здійснюють переважно проміжні, а власне кишкові — завершальні стадії гідролізу харчових речовин. Олігопептиди, що поступають в область щіткової облямівки, розщеплюються до амінокислот, здібних до всмоктування, за винятком гліцилгліцина і деяких діпептідов пролін, що містять, і оксипролін, які всмоктуються як такі. Дісахаріди, що поступають з їжею і переварювання крохмалю і глікогену, що утворюються в результаті, гидролізуются власне кишковими глікозідазамі до моносахаридів, які транспортуються через кишковий бар'єр у внутрішнє середовище організму. Трігліцеріди розщеплюються не лише під дією ліпази підшлункового соку, але і під впливом власне кишкового ферменту — моногліцерідліпази. Всмоктування відбувається в вигляді жирних кислот і b-моногліцерідов. Длінноцепочниє жирні кислоти в слизистій оболонці тонкої кишки знов естеріфіцируются і поступають в лімфу у вигляді хиломікронов (часток діаметром близько 0,5 мкм ) . жирні кислоти Короткоцепочниє не ресинтезіруются і поступають більшою мірою в кров, чим в лімфу. В цілому при мембранному П. розщеплюється велика частина всіх глікозідних і пептидних зв'язків і трігліцерідов. Мембранне П., на відміну від порожнинного, відбувається у стерильній зоні, т.к. мікроворсинки щіткової облямівки є своєрідним бактерійним фільтром, що відокремлює завершальні стадії гідролізу харчових речовин від заселеної бактеріями порожнини кишки. У нормі в процесах П. важливе значення мають мікроорганізми, а у деяких тварин — прості, населяючі різні відділи шлунково-кишкового тракту. Травні процеси в тонкій кишці розподілені неоднаково як в напрямі від її початку до кінця, так в е р б напрямі від крипт до верхівок ворсинок, що виражається у відповідній топографії кожного з травних ферментів, що здійснюють як порожнинне, так і мембранне П.
П. в товстих кишках практично відсутній. У їх вмісті виявляються незначні кількості ферментів і багата флора бактерій, що викликають зброджування вуглеводів і гниття білків, внаслідок чого утворюються органічні кислоти, гази (вуглекислий газ, метан і сірководень), отруйні речовини (фенол, скатол, індол, крезол), що знешкоджуються в печінці. Унаслідок мікробного бродіння розщеплюється клітковина. У товстих кишках переважають процеси зворотного всмоктування (реабсорбції) води, мінеральних і органічних компонентів харчової кашки — химуса . В товстих кишках всмоктуються до 95% води, а також електроліти, глюкоза, деякі вітаміни і амінокислоти, що продукуються мікробами кишкової флори . У міру просування і ущільнення вмісту кишечника формується кал, накопичення якого викликає акт дефекація .
Регуляція піщеваренія . Функції травної системи залежать від складу і кількості їжі, що вперше було підтверджене в експерименті І. П. Павловим . Існує певний зв'язок між вмістом різних травних ферментів і якістю їжі. В одних видів тварин (наприклад, в хижих) переважають протеолітичні ферменти, в інших (переважно рослиноїдних) — карбогідраза. Адаптивно-компенсаторні перебудови ферментних систем, що беруть участь в мембранному П., також обумовлені якістю їжі. Відмінності в наборі травних ферментів можуть бути як фенотипічні, так і генетичні походження. Наприклад, живлення може стимулювати не лише секрецію ферментів, але і їх синтез, а склад дієти може визначити співвідношення травних ферментів в даного організму. Якщо в травний канал поступають жири, білки і вуглеводи, в першу чергу перетравлюються жири, потім вуглеводи і, нарешті, білки. Діяльність травної системи координується за допомогою нервових і гуморальних регулювальників. Так, парасимпатична нервова система стимулює рухову функцію шлунково-кишкового тракту, а симпатична пригноблює її. Різні гормони, що особливо виробляються передньою долею гіпофіза і корою надниркових, впливають на синтез травних ферментів, їх перенесення і включення в ліпопротєїдниє комплекси мембрани мікроворсинок власне кишкових ферментів, на процеси всмоктування і моторику, а також секреторну функцію. Між виглядом їжі, тривалістю переварювання і швидкістю просування її по шлунково-кишковому тракту існує тонко збалансована залежність, здійснювана частково за допомогою місцевої регуляції, але в основному рефлекторно. У регуляції діяльності травної системи беруть участь сигнали, що поступають з рецепторів, локалізованих в більшості органів травного апарату і що забезпечують, зокрема, аналіз властивостей їжі в ротовій порожнині (див. Смак ). Значення відцентрової (еферентною) і доцентрової (аферентною) іннервації детально розглянуте при описі відповідних органів.
Літ.: Бабкин Би. П., Зовнішня секреція травних залоз, М.— Л., 1927; Павлов І. П., Лекції про роботу головних травних залоз, Полн. собр. соч.(вигадування), 2 видавництва, т. 2, кн. 2, М.— Л., 1951; Бабкин Би. П., Секреторний механізм травних залоз, Л., 1960; Проссер Л., Браун Ф., Порівняльна фізіологія тварин, пер.(переведення) з англ.(англійський), М., 1967; Угольов А. М. Травлення і його пристосовна еволюція, М., 1961; його ж, Мембранне травлення. Полісубстратниє процеси, організація і регуляція, Л., 1972; Bockus Н. L., Gastroenterology, v. 1—3, Phil.- L., 1963-65; Davenport Н. W., Physiology of the digestive tract, 2 ed., Chi., 1966; Handbook of physiology, sec. 6: Alimentary canal, v. 1—5, Wash., 1967—68; Jennings J. B., Feeding, digestion and assimilationin animals, 2 ed., L., 1972.