Нервова система, сукупність структур в організмі тварин і людини, об'єднуюча діяльність всіх органів і систем і що забезпечує функціонування організму як єдиного цілого в його постійній взаємодії із зовнішнім середовищем. Н. с. сприймає зовнішні і внутрішні роздратування, аналізує цю інформацію, відбирає і переробляє її і відповідно до цього регулює і координує функції організму.
Н. с. утворена головним чином нервовою тканиною, основний елемент якої — нервова клітина з відростками, що володіє високою збудливістю і здібністю до швидкого проведення збудження.
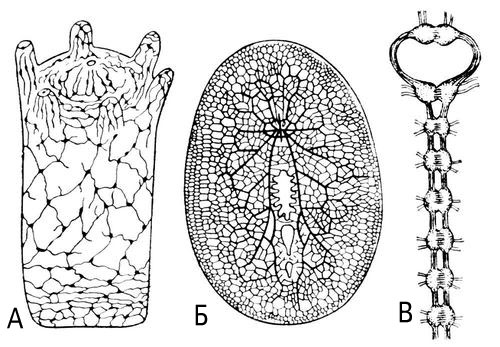
Еволюція Н. с., що зазнає нею в ході філогенезу, відрізняється великою складністю. В простих — одноклітинних організмів — Н. с. відсутній, але в деяких інфузорій є внутріклітинна сіточка, що виконує функцію проведення збудження до ін. елементам клітки. Найпримітивніша форма Н. с., що збереглася лише в нижчих кишечнополостних (гідра), — дифузна Н. с. ( мал. 1, А ) . Нервові клітини в кишечнополостних за допомогою відростків з'єднуються в мережу, в якій проведення збудження може здійснюватися на всіх напрямках. В процесі подальшої еволюції будова Н. с. ускладнюється. В що вільно живуть кишечнополостних відбувається освіта і занурення в глиб тіла скупчень нервових клітин — нервових вузлів (гангліїв), — зв'язки між якими встановлюються переважно за допомогою довгих відростків (нервових волокон, нервів). Поява такого дифузно-вузлового типа будови ( мал. 1, Би ) супроводиться розвитком спеціалізованих сприймаючих нервових структур (рецепторів ) що диференціюються відповідно до сприйманого ними вигляду енергії. Проведення збудження стає направленим. Наступний етап еволюції (кільчасті черв'яки, членистоногі, голкошкірі, молюски) — перехід до вузлового типа будови Н. с.: нервові клітини зосереджені у вузлах, зв'язаних нервовими волокнами між собою, а також з відповідними рецепторами і виконавськими органами. З'являється дістантная рецепція; серед нервових вузлів відбувається виділення домінуючих розташованих у тварин, що вільно пересуваються, на головному кінці тіла і пов'язаних з найбільш важливими дістантнимі рецепторами — органами чуття ( мал. 1, В ) . У зв'язку з тим, що головні вузли отримують при русі тварини найбільшу кількість інформації від зовнішнього світу, вони збільшуються, структура їх ускладнюється; тулубові ганглії наближаються до головних і зливаються з ними, утворюючи складні мозкові комплекси, які в якійсь мірі підпорядковують собі діяльність ін. вузлів.
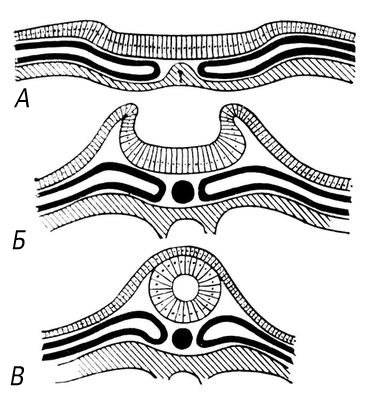
В хребетних тварин тип будови Н. с. різко відрізняється від вузлового типа, зазвичай властивого безхребетним. Центральна нервова система (ЦНС) представлена нервовою трубкою, розташованою на спинній стороні тіла, і складається із спинного і головного мозку. У ембріональному розвитку хребетних Н. с. утворюється із зовнішнього зародкового листка — ектодерми (спершу у вигляді нервової пластинки, що згортається в жолобок, а потім що перетворюється на нервову трубку) ( мал. 2 ). Зачаткові ектодермальні клітки диференціюються на нейробласти (клітки, що дають почало нейронам) і спонгиобласти (створюючі клітки нейроглії). З ектодермальних кліток, шляхом їх міграції, формуються і периферичні вузли, а сукупність відростків деяких нейробластов утворює черепномозкові і спинномозкові нерви, відношувані до периферичної Н. с. Головний кінець нервової трубки ділиться на 3 мозкових міхура — зачатки головного мозку . В процесі розвитку передній мозковий міхур розділяється на два, з яких один утворює кінцевий мозок, що включає великі півкулі і базальні ганглії, а другий — проміжний мозок. Середній мозковий міхур дає початок середньому мозку. Із заднього — утворюються мозочок, варолієв міст і довгастий мозок. Остання частина нервової трубки, зберігаючи трубчасту будову, утворює спинний мозок з потовщеннями в поперековій і плечовій областях. Як спинний, так і головний мозок хребетних покритий рядом оболонок і поміщений в кісткові покриви — череп і хребет.
В процесі еволюції відбувається подальше ускладнення структури Н. с. і удосконалення всіх форм її взаємодії із зовнішнім середовищем; при цьому всього більшого значення набувають передні відділи головного мозку, що прогресують в своєму розвитку. У риб передній мозок майже не диференційований, але у них добре розвинені задній, а також середній мозок; найбільшого розвитку у риб досягає мозочок. В земноводних і плазуючих задній мозок займає відносно менший об'єм, чим у риб, мозочок же поступається в розвитку середньому мозку, який ділиться на 2 частини, утворюючи двухолміє. Ускладнюється структурно і функціонально передній мозковий міхур, він диференціюється на проміжний мозок і 2 півкулі з розвиненою нервовою тканиною, створюючою так звану первинну кору мозку. Передній мозок, спочатку пов'язаний лише з нюхом, потім набуває і складніших функцій. Декілька відособлене місце в еволюційному ряду займають птиці, в яких домінують структури так званого мозкового ствола, тобто середній мозок і лише ті частини переднього мозку, які розташовані в глибині півкуль (базальні ганглії, проміжний мозок); сильно розвинений у птиць і мозочок; кора головного мозку диференційована слабо.
Вищого розвитку Н. с. досягає у ссавців. Головний кінець нервової трубки в ембріогенезі ділиться у них на 5 міхурів: передній — дає початок великим півкулям і проміжному мозку, середній — середньому мозку, задній — ділиться на власне задній (варолієв міст і мозочок) і довгастий мозок. Кора великих півкуль головного мозку утворює багаточисельні борозни і звивину. Первинна порожнина нервової трубки перетворюється на шлуночки мозку і спинномозковий канал. Нейронна організація мозку украй ускладнюється. Розвиток і диференціація структур Н. с. у високоорганізованих тварин зумовили її розділення на соматичну і вегетативну нервову систему . Особливість будови вегетативної Н. с. та, що її волокна, що відходять від ЦНС, не доходять безпосередньо до еффектора, а спочатку вступають в периферичні ганглії, де закінчуються на клітках, що віддають аксони вже безпосередньо на іннервіруємий орган. Залежно від того, де розташовані ганглії вегетативної Н. с., і деяких її функціональних особливостей вегетативну Н. с. ділять на 2 частини: парасимпатичну і симпатичну.
Структурна і функціональна одиниця Н. с. — нейрон, що складається з тіла нервової клітини і відростків — аксона і дендриту . Окрім нервових клітин, в структуру Н. с. входять гліальні клітки (див. Нейроглія ) . Нейрони є певною мірою самостійними одиницями — їх протоплазма не переходить з одного нейрона в іншій (див. Нейронна теорія ) . Взаємодія між нейронами здійснюється завдяки контактам між ними (див. Синапси ; мал. 3 ). В області контакту між закінченням одного нейрона і поверхнею іншого в більшості випадків зберігається особливий простір — синаптічеськая щілина — шириною в декілька сотів . Основні функції нейронів: сприйняття роздратувань, їх переробка, передача цієї інформації і формування у відповідь реакції. Залежно від типа і ходу нервових відростків (волокон), а також їх функцій нейрони підрозділяють на: а) рецепторні (аферентні), волокна яких проводять нервові імпульси від рецепторів в ЦНС; тіла їх знаходяться в спінальних гангліях або гангліях черепномозкових нервів; би) рухові (еферентні), зв'язуючі ЦНС з еффекторамі; тіла і дендрит їх знаходяться в ЦНС, а аксони виходять за її межі (за винятком еферентних нейронів вегетативної Н. с., тіла яких розташовані в периферичних гангліях); у) вставні (асоціативні) нейрони, службовці сполучними ланками між аферентними і еферентними нейронами; тіла і відростки їх розташовані в ЦНС.
Діяльність Н. с. грунтується на двох процесах: збудженні і гальмуванні . Збудження може бути таким, що поширюється (див. Імпульс нервовий ) або місцевим — що не поширюється, стаціонарним (останнє відкрито Н. Е. Введенським в 1901) Гальмування — процес, що тісно пов'язаний із збудженням і зовні виражається в зниженні збудливості кліток. Одна з характерних меж гальмівного процесу — відсутність здібності до активного поширення по нервових структурах (явище гальмування в нервових центрах вперше було встановлене І. М. Сеченовим в 1863).
Клітинні механізми збудження і гальмування детально вивчені. Тіло і відростки нервової клітини покриті мембраною, що постійно несе на собі різницю потенціалів (так званий мембранний потенціал). Роздратування розташованих на периферії чутливих закінчень аферентного нейрона перетвориться в зміну цій різниці потенціалів (див. Біоелектричні потенціали ) . нервовий імпульс, що Виникає внаслідок цього, поширюється по нервовому волокну і досягає його пресинаптичного закінчення, де викликає виділення в синаптічеськую щілину високоактивної хімічної речовини — медіатора . Під впливом останнього в постсинаптичній мембрані, чутливій до дії медіатора, відбувається молекулярна реорганізація поверхні. В результаті постсинаптична мембрана починає пропускати іони і деполяризується, унаслідок чого на ній виникає електрична реакція у вигляді місцевого збуджуючого постсинаптичного потенціалу (ВПСП), що знов генерує імпульс, що поширюється. Нервові імпульси, що виникають при збудженні особливих гальмівних нейронів, викликають гіперполяризацію постсинаптичної мембрани і, відповідно, гальмівний постсинаптичний потенціал (ТПСП). Окрім цього, встановлений і ін. вигляд гальмування, що формується в пресинаптичній структурі, — пресинаптичне гальмування, що обумовлює тривале зниження ефективності синаптічеськой передачі (див. Мембранна теорія збудження ) .
В основі діяльності Н. с. лежить рефлекс, тобто реакція організму на роздратування рецепторів, здійснювана за посередництва Н. с. Термін «рефлекс» був вперше введений у фізіологію Р. , що зароджувалася, Декартом в 1649, хоча конкретних уявлень про те, як здійснюється рефлекторна діяльність, у той час ще не було. Такі відомості були отримані лише значно пізніше, коли морфологи приступили до дослідження будови і функцій нервових клітин (Р. Дютроше 1824; До. Еренберг, 1836; Я. Пуркине, 1837; До. Гольджі, 1873; С. Рамон-і-Кахаль, 1909), а фізіологами були вивчені основні властивості нервової тканини (Л. Гальвані, 1791; До. Маттеуччи, 1847; Е. Дюбуа-Реймон, 1848—49; Н. Е. Введенський, 1901; А. Ф. Самойлов, 1924; Д. С. Воронцов, 1924; і ін.). В кінці 19 і початку 20 вв.(століття) були створені карти розташування нервових центрів і нервових доріг в мозку, а також отримані відомості про основні рефлекторні процеси і про локалізацію функцій у мозку, з тих пір постійно поповнювані і розширювані (І. М. Сеченов, 1863; Н. А. Міславський, 1885; Ст М. Бехтерев, 1903; І. П. Павлов, 1903; Ч. Шеррінгтон, 1906; А. А. Ухтомський, 1911; І. С. Беріташвілі, 1930; Л. А. Орбелі, 1932; Дж. Фултон, 1932; Е. Едріан, 1932; П. До. Анохин, 1935; До. М. Биків, 1941; Х. Мегоун, 1946; і ін.). Всі рефлекторні процеси пов'язані з поширенням збудження по певних нервових структурах — рефлекторним дугам . Основні елементи рефлекторної дуги: рецептори, доцентрова (аферентний) нервова дорога, внутрішньоцентральні структури різної складності, відцентрова (еферентний) нервова дорога і виконавський орган (еффектор). Різні групи рецепторів збуджуються подразниками різної модальності (тобто якісній специфічності) і сприймають роздратування, витікаючі як із зовнішнього середовища (екстерорецепції — органи зору, слуху нюх і ін.), так і з внутрішнього середовища організму (інтерорецептори, що збуджуються при механічних, хімічних, температурних і ін. роздратуваннях внутрішніх органів, м'язів і ін.). Нервові сигнали, що несуть в ЦНС інформацію від рецепторів по нервових волокнах, позбавлені модальності і зазвичай передаються у вигляді серії однорідних імпульсів. Інформація про різні характеристики роздратувань кодується змінами частоти імпульсів, а також пріуроченностью нервовою імпульсациі до певним волокнам (так зване просторово-часове кодування).
Сукупність рецепторів даної області тіла тварини або людини, роздратування яких викликає певний тип рефлекторної реакції, називається рецептивним полем рефлексу. Такі поля можуть накладатися один на одного. Сукупність нервових утворень, зосереджених в ЦНС і відповідальних за здійснення даного рефлекторного акту, позначають терміном нервовий центр . На окремому нейроні в Н. с. може сходитися величезне число закінчень волокон, що несуть імпульси від ін. нервових клітин. У кожен даний момент в результаті складної синаптічеськой переробки цього потоку імпульсів забезпечується подальше проведення лише одного, певного сигналу — принцип конвергенції, лежачий в основі діяльності всіх рівнів Н. с. («принцип кінцевої загальної дороги» Шеррінгтона що отримав розвиток в працях Ухтомського і ін.).
Просторово-часова суммация синаптічеських процесів служить основою для різних форм виборчого функціонального об'єднання нервових клітин, лежачого в основі аналізу що поступає в Н. с. інформації і вироблення потім команд для виконання різних у відповідь реакцій організму. Такі команди, як і аферентні сигнали, передаються від однієї клітки до іншої і від ЦНС до виконавських органів в вигляді послідовностей нервових імпульсів, що виникають в клітці у тому випадку, коли збуджуючі і гальмівні синаптічеськие процеси, що підсумовуються, досягають певного (критичного для даної клітки) рівня — порогу збудження.
Не дивлячись на спадково закріплений характер зв'язків в основних рефлекторних дугах, характер рефлекторної реакції може в значній мірі змінюватися залежно від стану центральних утворень, через які вони здійснюються. Так, різке підвищення або пониження збудливості центральних структур рефлекторної дуги може не лише кількісно змінити реакцію, але і привести до певних якісних змін в характері рефлексу. Прикладом такої зміни може служити явище домінанти .
Важливе значення для нормального протікання рефлекторної діяльності має механізм так званої зворотною афферентациі — інформації про результат виконання даної рефлекторної реакції, що поступає по аферентних дорогах від виконавських органів. На підставі цих відомостей у випадку, якщо результат незадовільний, у функціональній системі, що сформувалася, можуть відбуватися перебудови діяльності окремих елементів до тих пір, поки результат не стане відповідати рівню, необхідному для організму (П. До. Анохин, 1935).
Всю сукупність рефлекторних реакцій організму ділять на дві основні групи: безумовні рефлекси — природжені, здійснювані по спадково закріплених нервових дорогах, і умовні рефлекси, придбані протягом індивідуального життя організму шляхом освіти в ЦНС тимчасових зв'язків. Здатність утворення таких зв'язків властива лише вищому для даного вигляду тварин відділу Н. с. (для ссавців і людини — це кора головного мозку). Утворення умовнорефлекторних зв'язків дозволяє організму найбільш абсолютно і тонко пристосовуватися до умов існування, що постійно змінюються. Умовні рефлекси були відкриті і вивчені І. П. Павловим в кінці 19 — початку 20 вв.(століття) Дослідження умовнорефлекторної діяльності тварин і людини привело його до створення учення про вищій нервовій діяльності (ВНД) і аналізаторах. Кожен аналізатор складається із сприймаючої частини — рецептора, провідних доріг і аналізуючих структур ЦНС, що обов'язково включають її вищий відділ. Кора головного мозку у вищих тварин — сукупність кіркових кінців аналізаторів; вона здійснює вищі форми діяльності аналізатора і інтеграційної, забезпечуючи досконалі і якнайтонші форми взаємодії організму із зовнішнім середовищем.
Н. с. володіє здатністю не лише негайно переробляти інформацію, що поступає в неї, за допомогою механізму взаємодіючих синаптічеських процесів, але і зберігати сліди минулої активності (механізми пам'яті ) . Клітинні механізми збереження у вищих відділах Н. с. тривалих слідів нервових процесів, лежачі в основі пам'яті, інтенсивно вивчаються.
Поряд з перерахованими вище функціями Н. с. здійснює також регулюючі впливи на обмінні процеси в тканинах — адаптаційно-трофічну функцію (І. П. Павлов, Л. А. Орбелі, А. В. Тонких і ін.). При перерізанні або пошкодженні нервових волокон властивості іннервіруємих ними кліток змінюються (це торкається як физико-хімічних властивостей поверхневої мембрани, так і біохімічних процесів в протоплазмі), що, у свою чергу, супроводиться глибокими порушеннями в стані органів і тканин (наприклад, трофічними виразками). Якщо іннервація відновлюється (у зв'язку з регенерацією нервових волокон), то вказані порушення можуть зникнути.
Літ.: Орбелі Л. А., Лекції з фізіології нервової системи, 3 видавництва, М. — Л., 1938; його ж, Ізбр. праці, т. 1—5, М. — Л., 1961—68; Ухтомський А. А., Собр. соч.(вигадування), т. 1—6, Л., 1945—62; Павлов І. П., Полн. собр. соч.(вигадування), 2 видавництва, т. 2, М., 1951; Сеченов І. М., Ізбр. проїзв.(твір), т. 1, [М.], 1952; Коштоянц Х. С., Основи порівняй тільній фізіології, т. 2, М., 1957; Беріташвілі І. С., Загальна фізіологія м'язової і нервової системи, 3 видавництва, т. 1, М., 1959; Сепп Е. До., Історія розвитку нервової системи хребетних, 2 видавництва, М., 1959: Екклс Дж., Фізіологія нервових клітин, пер.(переведення) з англ.(англійський) М., 1959; Беклемішев Ст Н., Основи порівняльної анатомії безхребетних, 3 видавництва, т. 2, М., 1964; Катц Би., Нерв, м'яз і синапс, пер.(переведення) з англ.(англійський), М., 1968; Окс С., Основи нейрофізіології, пер.(переведення) з англ.(англійський), М., 1969; Шеррінгтон Ч., Інтеграційна діяльність нервової системи, пер.(переведення) з англ.(англійський), Л., 1969: Костюк П. Р., Фізіологія центральної нервової системи, До., 1971; Ariens Kappers С. U., Huber G. С., Crosby E. С., The comparative anatomy of the nervous system of vertebrates, including man, v. 1—2, N. Y., 1936; Bullock T. Н., Horridge G. A., Structure and function in the nervous systems of invertebrates, v. 1—2, S. F. — L., 1965.