Біоелектричні потенціали, електричні потенціали, що виникають в тканинах і окремих клітках людини, тварин і рослин, найважливіші компоненти процесів збудження і гальмування . Дослідження Б. п. має велике значення для розуміння физико-хімічних і фізіологічних процесів в живих системах і застосовується в клініці з діагностичною метою (електрокардіографія,електроенцефалографія,електроміографія і ін.).
Перші дані про існування Б. п. («тваринної електрики») були отримані в 3-ій чверті 18 ст при вивченні природи «удару», що наноситься деякими рибами з електричними органами при захисті або нападі. До цього ж часу відноситься початок досліджень італійського фізіолога і лікарки Л. Гальвані, що заклали основу учення о Б. п. Багатолітня наукова суперечка (1791—97) між Л. Гальвані і фізиком А. Вольта про природу «тваринної електрики» завершилася двома крупними відкриттями: були отримані факти про існування біоелектричних явищ в живих тканинах і відкритий новий принцип здобуття електричного струму за допомогою різнорідних металів — створений гальванічний елемент (вольтів стовп ). Правильна оцінка спостережень Гальвані стала можливою лише після вживання досить чутливих приладів електровимірювань — гальванометрів . Перші такі дослідження були проведені італійським фізиком К. Маттеуччи (1837). Систематичне вивчення Б. п. було почато німецьким фізіологом Е . Дюбуа-Реймоном (1848), який довів існування Б. п. в нервах і м'язах у спокої і при збудженні. Але йому не удалося (через велику інерційність гальванометра) зареєструвати швидкі, триваючі тисячні долі сік коливання Б. п. при проведенні імпульсів уздовж нервів і м'язів. У 1886 німецький фізіолог Ю. Бернштейн проаналізував форму потенціалу дії; французький учений Е. Ж. Марей (1875) застосував для запису коливань потенціалів серця, що б'ється, капілярну електрометрію; російський фізіолог Н. Е. Введенський використовував (1883) для прослухування ритмічних розрядів імпульсів в нерві і м'язі телефон, а голландський фізіолог В. Ейнтховен (1903) ввів в експеримент і клінічну практику струнний гальванометр — високочутливий і малоінерційний прилад для реєстрації електричних струмів в тканинах. Значний вклад у вивчення Б. п. внесли російські фізіологи: В. В. Правдіч-Немінський (1913—21) вперше зареєстрував електроенцефалограму, А. Ф. Самойлов (1929) досліджував природу нервово-м'язової передачі збудження, а Д. С. Воронцов (1932) відкрив коливання слідів Би. п., супроводжуючі потенціал дії в нервових волокнах. Подальший прогрес у вивченні Б. п. був тісно пов'язаний з успіхами електроніки, що дозволили застосувати у фізіологічному експерименті електронні підсилювачі і осцилографи (роботи американських фізіологів Р. Бішопа, Дж. Ерлангера і Г. Гассера в 30—40-х рр. 20в.). Вивчення Б.п. у окремих клітках і волокнах стало можливим з розробкою мікроелектродної техніки . Важливе значення для з'ясування механізмів генерації Б. п. мало використання гігантських нервових волокон головоногих молюсків, головним чином кальмара. Діаметр цих волокон в 50 — 100 разів більший, ніж у хребетних тварин, він досягає 0,5—1 мм , що дозволяє вводити всередину волокна мікроелектроди, ін'єктувати в протоплазму різні речовини і т.п. Вивчення іонної проникності мембрани гігантських нервових волокон дозволило англійським фізіологам А. Ходжкину, А. Хакслі і Б. Катцу (1947—52) сформулювати сучасну мембранну теорію збудження .
Розрізняють наступні основні види Б. п. нервових і м'язових клітин: потенціал спокою, потенціал дії, збуджуючі і гальмівні постсинаптичні потенціали, генераторні потенціали.
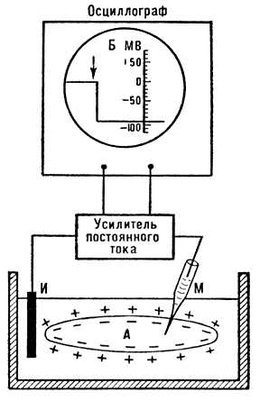
Потенціал спокою (ПП, мембранний потенціал спокою). В живих клітин у спокої між внутрішнім вмістом клітки і зовнішнім розчином існує різниця потенціалів (ПП) порядку 60— 90мв , яка локалізована на поверхневій мембрані. Внутрішня сторона мембрани заряджена електронегативно по відношенню до зовнішньої ( мал. 1 ). ПП обумовлений виборчою проникністю мембрани, що покоїться, для іонів К + (Ю. Бернштейн, 1902, 1912; А. Ходжкин і Б. Катц, 1947). Концентрація К + в протоплазмі приблизно в 50 разів вище, ніж в позаклітинній рідині, тому дифундує з клітки, іони виносять на зовнішню сторону мембрани позитивні заряди, при цьому внутрішня сторона мембрани, практично не проникної для крупних органічних аніонів, набуває негативного потенціалу. Оскільки проникність мембрани у спокої для Na + приблизно в 100 разів нижче, ніж для К + , дифузія натрію з позаклітинної рідини (де він є основним катіоном) в протоплазму мала і лише трохи знижує ПП обумовлений іонами К + . У скелетних м'язових волокнах у виникненні потенціалу спокою важливу роль грають також іони Cl - , дифундуючі всередину клітки. Наслідком ПП є струм спокою, що реєструється між пошкодженою і інтактною ділянками нерва або м'яза при додатку електродів, що відводять. Мембрани нервових і м'язових клітин (волокон) здатні змінювати іонну проникність у відповідь на зрушення мембранного потенціалу. При збільшенні ПП (гіперполяризація мембрани) проникність поверхневих клітинних мембран для Na + і К + падає, а при зменшенні ПП (деполяризація) вона зростає, причому швидкість змін проникності для Na + значно перевищує швидкість збільшення проникності мембрани для К + .
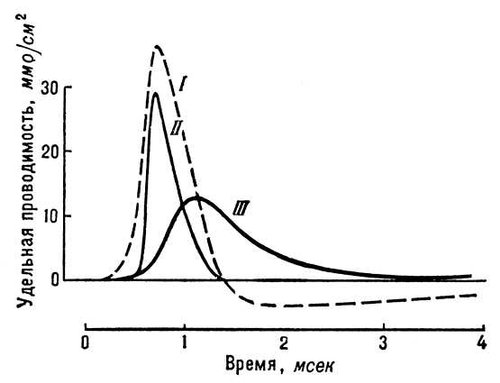
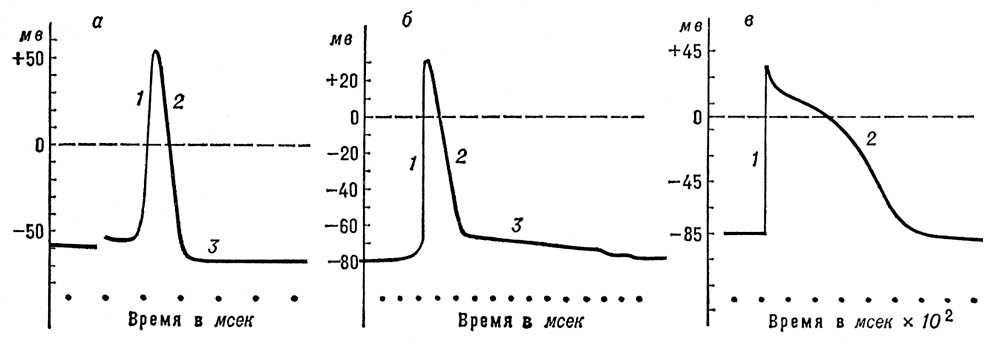
Потенціал дії (ПД). Всі подразники, що діють на клітку, викликають в першу черга зниження ПП; коли воно досягає критичного значення (порогу), виникає активна відповідь, що поширюється, — ПД ( мал. 2 ). Під час висхідної фази ПД короткочасно перекручується потенціал на мембрані: її внутрішня сторона, заряджена у спокої електронегативно, набуває в цей час позитивного потенціалу. Досягнувши вершини, ПД починає падати (низхідна фаза ПД), і потенціал на мембрані повертається до рівня, близького до початкового, — Пп. Повне відновлення ПП відбувається лише після закінчення коливань слідів потенціалу — деполяризації сліду або гіперполяризації, тривалість яких зазвичай значно перевершує тривалість піку ПД. Згідно мембранної теорії, деполяризація мембрани, викликана дією подразника, приводить до посилення потоку Na + всередину клітки, що зменшує негативний потенціал внутрішньої сторони мембрани — підсилює її деполяризацію. Це, у свою чергу викликає подальше підвищення проникності для Na + і нове посилення деполяризації і т.д. В результаті такого вибухового кругового процесу, т.з. регенеративній деполяризації, відбувається збочення мембранного потенціалу, характерне для ПД. Підвищення проникності для Na + дуже короткочасно і змінявся її падінням ( мал. 3 ), а отже, зменшенням потоку Na + всередину клітки. Проникність для К + , на відміну від проникності для Na + , продовжує збільшуватися, що приводить до посилення потоку К + з клітки. В результаті цих змін ПД починає падати, що веде до відновлення Пп. Такий механізм генерації ПД в більшості збудливих тканин. Існують, проте, клітки (м'язові волокна ракоподібних, нервові клітини в ряду брюхоногих молюсків, деякі рослинні клітки), в яких висхідна фаза ПД обумовлена підвищенням проникність мембрани не для іонів Na + , а для іонів Ca + . Своєрідний також механізм генерації ПД в м'язових волокнах серця, для яких характерне тривале плато на низхідній фазі ПД ( мал. 2 , би). Нерівність концентрацій іонів К + і Na + (або Ca + ) усередині і зовні клітки (волокна) підтримується спеціальним механізмом (т.з. «натрієвим насосом» ), що виштовхує іони Na + з клітки і що нагнітає іони К + в протоплазму, що вимагає витрати енергії, яка черпається кліткою в процесах обміну речовин.
Амплітуда ПД більшості нервових і м'язових волокон приблизно однакова: 110—120 мв. Тривалість ПД варіює в широких межах: у теплокровних тварин тривалість ПД нервових волокон, найшвидше провідних збудження, — 0,3—0,4 мсек , у волокон же м'язів серця — 50—600 мсек. В рослинних клітках прісноводої водорості хара ПД продовжується близько 20 сек. Характерною особливістю ПД, що відрізняє його від інших форм відповіді клітки на роздратування, є те, що він підкоряється правилу «все або нічого», тобто виникає лише при досягненні подразником деякого порогового значення, і подальше збільшення інтенсивності подразника вже не позначається ні на амплітуді, ні на тривалості ПД. Потенціал дії — один з найважливіших компонентів процесу збудження. У нервових волокнах він забезпечує проведення збудження від чутливих закінчень (рецепторів ) до тіла нервової клітини і від неї — до синаптічеським закінчень (див. Синапси ), розташованих на різних нервових, мишечних або залізистих клітках. Поступаючи в еффекторниє закінчення, ПД викликає виділення (секрецію) певній порції специфічних хімічних речовин, т.з. медіаторів, що роблять збуджуючий або гальмівний вплив на відповідні клітки. У м'язових волокнах ПД, що поширюється, викликає ланцюг физико-хімічних реакцій, лежачих в основі процесу скорочення м'язів. Проведення ПД уздовж нервових і м'язових волокон здійснюється т.з. локальними струмами, або струмами дії, що виникають між збудженим (деполяризованим) і сусідніми з ним ділянками мембрани, що покояться (див. Збудження ). Струми дії реєструються звичайними позаклітинними електродами; при цьому крива має двофазний характер: перша фаза відповідає приходу ПД під ближній електрод, друга — під далекий електрод ( мал. 4 ).
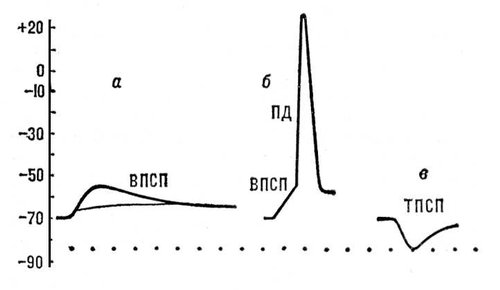
Постсинаптичні потенціали (ПСП) виникають в ділянках мембрани нервових або м'язових клітин, що безпосередньо граничать з синаптічеськимі закінченнями. Вони мають амплітуду порядка декілька мв і тривалість 10—15 мсек. ПСП підрозділяються на збуджуючих (ВПСП) і гальмівних (ТПСП). ВПСП є місцевою деполяризацією постсинаптичної мембрани, обумовленою дією відповідного медіатора (наприклад, ацетілхоліну в нервово-м'язовому з'єднанні). Досягши ВПСП деякого порогового (критичного) значення в клітці виникає ПД, що поширюється ( мал. 5 , а, би). ТПСП виражається місцевою гіперполяризацією мембрани, обумовленої дією гальмівного медіатора ( мал. 5 , в). На відміну від ПД, амплітуда ПСП поступово збільшується із збільшенням кількості медіатора, що виділився з нервового закінчення. ВПСП і ТПСП підсумовуються один з одним при одночасному або послідовному вступі нервових імпульсів до закінчень, розташованих на мембрані однієї і тієї ж клітки.
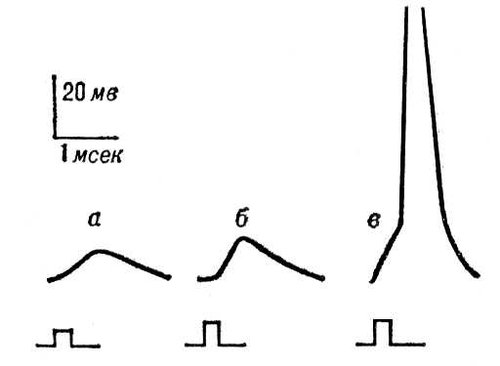
Генераторні потенціали виникають в мембрані чутливих нервових закінчень — рецепторів. Вони зовні схожі з ВПСП — їх амплітуда порядка декілька мв і залежить від сили прикладеного до рецептора роздратування ( мал. 6 ). Коли генераторний потенціал досягає порогового (критичного) значення, в сусідній ділянці мембрани нервового волокна виникає ПД, що поширюється. Іонний механізм генераторних потенціалів ще недостатньо вивчений.
Поряд з перерахованими що відносно швидко розвиваються Б. п., в нервових клітинах, волокнах гладких м'язів і деяких рослинних клітках реєструються також дуже повільні коливання мембранного потенціалу невідомої природи, причому на гребені хвилі деполяризації мембрани часто виникають розряди імпульсів.
Все Б. п. можуть бути зареєстровані і точно виміряні лише за допомогою внутріклітинних мікроелектродів, що дозволяють відводити різниці потенціалів між внутрішньою і зовнішньою сторонами мембрани клітки. При відведенні коливань Би. п. від цілих нервів, м'язів або мозку за допомогою поверхневих електродів реєструється лише сумарно потенціал безлічі синхронно або, частіше асихронно працюючих кліток. Так, електроміограма є результатом складання (інтерференції) ПД безлічі скелетних м'язових волокон; електрокардіограма — результуюча коливань електричних потенціалів м'язових волокон різних відділів серця; електроенцефалограма — результат суммациі головним чином ВПСП і ТПСП безлічі кліток різних шарів кори великих півкуль. Реєстрація таких інтерференційних електрограм, хоча і не дозволяє аналізувати коливання Б. п. окремих кліток, має важливе значення для думки про стан досліджуваного органу в цілому. У клінічній практиці електроміограму, електрокардіограму і електроенцефалограму реєструють за допомогою електродів, розташованих на шкірі відповідних частин тіла. Оцінка даних, отриманих цими методами, заснована на зіставленні змін характеру відповідній кривій з результатами клінічних, фізіологічних і патологоанатомічних досліджень.
Літ.; Фізіологія людини, М., 1966; Гальвані Л. і Вольта А., Ізбр. роботи про тваринну електрику, М. — Л., 1937; Ходжкин А., Нервовий імпульс, пер.(переведення) з англ.(англійський), М., 1965; Екклс Дж., Фізіологія нервових клітин, пер.(переведення) з англ.(англійський), М., 1959; його ж, Фізіологія синапсів, М., 1966; Катц Би., Нерв, м'яз і синапс, пер.(переведення) з англ.(англійський), М., 1968; Ходоров Би. І., Проблема збудливості, Л., 1969.