Синапси (від греч.(грецький) sýnapsis — з'єднання, зв'язок), спеціалізовані функціональні контакти між збудливими клітками, службовці для передачі і перетворення сигналів. Термін «З.» був вперше використаний англійським фізіологом Ч. Шеррінгтоном в 1897 для позначення контактів між нейронами. Контакти між аксонами нейронів і клітками виконавських органів часто визначають як з'єднання, хоча вони представляють різновид С. Поськольку С. — єдина дорога, за допомогою якої нейрони можуть повідомлятися один з одним, вони забезпечують всі основні прояви активності нервової системи і інтеграційну діяльність мозку. У С. входять пресинаптична частина (синаптічеськоє закінчення), синаптічеськая щілина (що розділяє 2 клітки) і постсинаптична частина (ділянка клітки, до якого прілежіт синаптічеськоє закінчення).
Міжнейронні С. в більшості випадків утворені закінченнями аксонів одних нервових клітин і тілом, дендритом або аксонами інших. Відповідно до цього розрізняють аксо-соматічні, аксо-дендрітні і аксо-аксонниє С. Ввіду того що поверхня дендриту переважає, найбільш багаточисельні аксо-дендрітні С. Число синаптічеських контактів на різних нейронах центральної нервової системи варіює в широких межах. На одних клітках закінчуються сотні або тисячі окремих пресинаптичних волокон, інші нейрони мають єдиний С. Крупний нейрон ретикулярної формації ствола мозку отримує понад 4000 синаптічеських контактів; на деяких клітках зразкове число синаптічеських контактів складає більше 10000—20000. Щільність розташування С. на поверхні нейрона може досягати 15—20 на 100 мкм 2 .
По функціональному значенню С. можуть бути збуджуючими і гальмівними відповідно до того, активують вони або пригнічують діяльність відповідної клітки. У тому і в іншому випадку передача через С. може здійснюватися за допомогою хімічного або електричного механізму. Крім того, існують змішані С., поєднуючі хімічні і електричні механізми передачі. Поширеніші С. з хімічним механізмом. У них сигнал з пресинаптичної мембрани передається на постсинаптичну за допомогою медіатора — хімічні сполуки, молекули якого здатні реагувати із специфічними рецепторами постсинаптичної мембрани і змінювати її проникність до іонів, викликаючи генерацію місцевого, нерегенеративного потенціалу. У електричних С. струм з активованої пресинаптичної мембрани безпосередньо впливає на постсинаптичну мембрану.
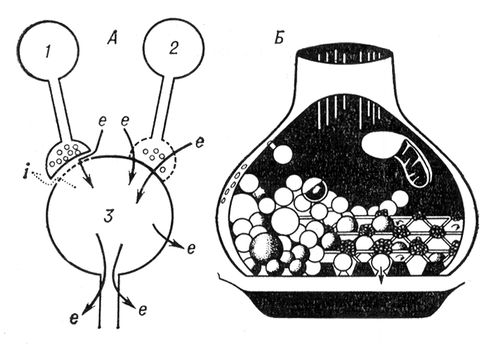
С. з хімічними і електричними механізмами передачі характеризуються специфічними структурними особливостями. У першому типові С. пресинаптичне закінчення містить т.з. синаптічеськие бульбашки, або везикули, що містять високі концентрації медіатора. Пре- і постсинаптичні мембрани розділені синаптічеськой щілиною, ширина якої зазвичай складає 150—200 Å, а в деяких С. досягає 1000 і більш Å. Синаптічеськие бульбашки мають тенденцію концентруватися в внутрішній поверхні пресинаптичні мембрани, протистоячій синаптічеськой щілині. Вони можуть виходити з пресинаптичного закінчення в місцях перерви мембрани, проникати в синаптічеськую щілину і контактувати з постсинаптичною мембраною. Розташування синаптічеських бульбашок і їх кількість змінюються в результаті нервової активності. Для постсинаптичної мембрани в хімічних С. характерні потовщення, на яких можна виділити особливі активні зони, мабуть, зв'язані з хеморецепторной спеціалізацією мембрани. У електричних С. щілина між пре- і постсинаптичними мембранами відсутня і інколи спостерігається їх повне злиття. Схематично обидва типи С. показані на. демонструє пресинаптичне закінчення хімічних С. з упакованими в нім пресинаптичними бульбашками. Процес передачі збуджуючих або гальмівних ефектів в С. з хімічним механізмом зводиться до наступних процесів: нервовий імпульс, що приходить в пресинаптичне закінчення викликає деполяризацію пресинаптичної мембрани, що у свою чергу збільшує її проникність до іонів кальцію. Входження іонів кальцію всередину пресинаптичного закінчення викликає звільнення медіатора, який дифундує через синаптічеськую щілину і реагує з рецепторами постсинаптичної мембрани.
Ця реакція зазвичай приводить до збільшення проникності постсинаптичної мембрани до одному або декількох іонів і генерації потенціалу постсинаптичного . В разі збуджуючих С. збільшується натрієва провідність, інколи паралельно з калієвою провідністю, що приводить до деполяризації і збудження постсинаптичної клітки. У гальмівних С. збільшується проникність постсинаптичної мембрани до іонів хлору, а інколи паралельно до іонів калія. Цей ефект зазвичай супроводиться гіперполяризацією. Найбільш важливе значення для здійснення синаптічеського гальмування має саме збільшення провідності постсинаптичної мембрани, яке шунтує збуджуючі ефекти. Медіатор може впливати також на метаболічні процеси постсинаптичного нейрона, викликаючи тривалі постсинаптичні потенціали. У С. з електричним механізмом струми дії пресинаптичного закінчення прямо впливають на постсинаптичну клітку без участі проміжної хімічної ланки унаслідок майже повної відсутності синаптічеськой щілини (її ширина не перевищує 20 Å). Це усуває шунтування струму, поточного від пресинаптичної клітки до постсинаптичної. Імпульс, що генерується в пресинаптичній мембрані, передається на постсинаптичну мембрану пасивно, електротонічеськи, як по кабельних структурах ( мал. 2 ).
Особливість електротонічних С. — існування каналів, низькомолекулярних з'єднань, що дозволяють молекулам, проходіть з цитоплазми однієї клітки в цитоплазму інший. Ці канали не сполучаються з позаклітинним простором і відсутні в інших ділянках мембрани. Велика частина нервових процесів може здійснюватися за допомогою як хімічних, так і електротонічних С. Електротонічеськие С. забезпечують прудкість і стабільність передачі, менш чутливі до коливань температури. Хімічний механізм дозволяє змінювати ефективність С. в результаті попередньої активності, надійніше забезпечує однобічність проведення.
Літ.: Екклс Дж., Фізіологія синапсів, пер.(переведення) з англ.(англійський), М., 1966; Катц Би., Нерв, м'яз і синапс, пер.(переведення) з англ.(англійський), М., 1968; Акерт До., Порівняння рухових кінцевих пластинок і центральних синапсів. Ультраструктурноє дослідження, «Журнал еволюційної біохімії і фізіології», 1975 т. 11 №2; De Robertís Е. D., Histophysíology of synapses and neurosecretion, Oxf., 1964; Structure and function of synapses, ed. G. D. Pappas, D. P. Purpura, N. Y., 1972; Shapovalov A. I., Neuronal organization and synaptic mechanisms of supraspinal motor control in vertebrates, «Rev. Physiol., Biochem., PHARMACOL», 1975, v. 72.