Кора великих півкуль головного мозку, шар сірої речовини товщиною 1—5 мм, що покриває півкулі великого мозку ссавців тварин і людини. Ця частина головного мозку, що розвинулася на пізніх етапах еволюції тваринного світу, грає виключно важливу роль в здійсненні психічною, або вищою нервовою діяльності, хоча ця діяльність є результатом роботи мозку як єдиного цілого. Завдяки двостороннім зв'язкам з відділами нервової системи, що пролягають нижче, кора може брати участь в регуляції і координації всіх функцій організму. У людини кора складає в середньому 44% від об'єму всієї півкулі в цілому. Її поверхня досягає 1468—1670 см 2 .
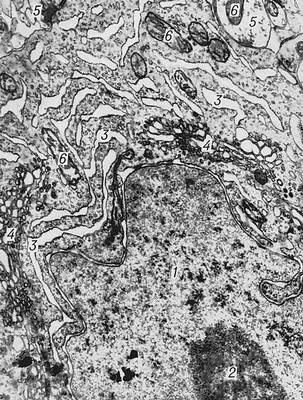
Будова кори. Характерною особливістю будови кори є орієнтований, горизонтально-вертикальний розподіл складових її нервових клітин по шарах і колонках; таким чином, кіркова структура відрізняється просторово впорядкованим розташуванням функціонуючих одиниць і зв'язків між ними ( мал. 1 ) . Простір між тілами і відростками нервових клітин кори заповнений нейроглією і судинною мережею (капілярами). Нейрони кори підрозділяються на 3 основних типа: пірамідні (80—90% всіх кліток кори), зірчасті і веретеноподібні. Основні функціональний елемент кори — аферентно-еферентний (тобто що сприймає доцентрові і що посилає відцентрові стимул-реакції) длінноаксонний пірамідний нейрон ( мал. 2 ) . Зірчасті клітки відрізняються слабким розвитком дендриту і потужним розвитком аксонів, які не виходять за межі поперечника кори і охоплюють своїми розгалуженнями групи пірамідних кліток. Зірчасті клітки виконують роль сприймаючих і синхронізуючих елементів, здатних координувати (одночасно гальмувати або збуджувати) просторово близькі групи пірамідних нейронів. Кірковий нейрон характеризується складною субмікроскопічною будовою (див. Клітка ) . Різні по топографії ділянки кори відрізняються щільністю розташування кліток, їх величиною і іншими характеристиками пошарової і колончатої структури. Всі ці показники визначають архітектуру кори, або її цитоархитектонику ( див. мал.(малюнок) 1 і 3 ) .
Найбільш крупні підрозділи території кори — древня (палеокортекс), стара (архикортекс), нова (неокортекс) і проміжна кора. Поверхня нової кори у людини займає 95,6%, старою 2,2%, древньою 0,6%, проміжною 1,6%.
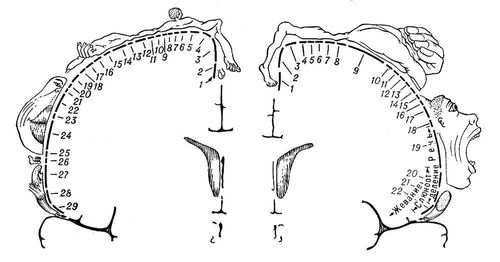
Якщо уявити собі кору мозку у вигляді єдиного покриву (плаща), що одягає поверхню півкуль, то основна центральна частина його складе нова кора, тоді як древня, стара і проміжна займуть місце на периферії, тобто по краях цього плаща. Древня кора у людини і вищих ссавців складається з одного клітинного шару, нечітко відокремленого від підкіркових ядер, що пролягають нижче; стара кора повністю відокремлена від останніх і представлена 2—3 шарами; нова кора полягає, як правило, з 6—7 шарів кліток; проміжні формації — перехідні структури між полями старої і нової кори, а також древньої і нової кори — з 4—5 шарів кліток. Неокортекс підрозділяється на наступні області: прецентральную, центральну для поста, скроневу, ніжнетеменную, верхнетеменную, скронево-тім'яно-потиличну, потиличну, островковую і лімбічеськую. У свою чергу, області підрозділяються на підобласті і поля. Основний тип прямих і зворотних зв'язків нової кори — вертикальні пучки волокон, що приносять інформацію з підкіркових структур до кори і що посилають її від кори в ці ж підкіркові утворення. Поряд з вертикальними зв'язками є внутрікортікальниє — горизонтальні — пучки асоціативних волокон, проходящие на різних рівнях кора і в білій речовині під корою. Горизонтальні пучки найбільш характерні для I і III шарів кори, а в деяких полях для V шаруючи. Горизонтальні пучки забезпечують обмін інформацією як між полями, розташованими на сусідній звивині, так і між віддаленими ділянками кори (наприклад, лобовою і потиличною).
Функціональні особливості кори обумовлюються згаданим вище розподілом нервових клітин і їх зв'язків по шарах і колонках. На кіркові нейрони можлива конвергенція (сходження) імпульсів від різних органів чуття. Згідно з сучасними виставами, подібна конвергенція різнорідних збуджень — нейрофізіологічний механізм інтеграційної діяльності головного мозку, тобто аналізу і синтезу у відповідь діяльності організму. Істотне значення має і те, що нейрони зведені в комплекси, мабуть реалізовуючі результати конвергенції збуджень на окремі нейрони. Одна з основних морфо-функціональніх одиниць кори — комплекс, званий колонкою кліток, який проходить через всі кіркові шари і складається з кліток, розташованих на одному перпендикулярі до поверхні кори. Клітки в колонці тісно зв'язані між собою і отримують загальну аферентну гілочку з підкірки. Кожна колонка кліток відповідає за сприйняття переважно одного вигляду чутливості. Наприклад, якщо у кірковому кінці шкірного аналізатора одна з колонок реагує на дотик до шкіри, то інша — на рух кінцівки в суглобі. У зоровому аналізаторі функції сприйняття зорових образів також розподілені по колонках. Наприклад, одна з колонок сприймає рух предмету в горизонтальній плоскості, сусідня — в вертикальною і т. п.
Другий комплекс кліток нової кори — шар — орієнтований в горизонтальній плоскості. Вважають, що дрібноклітинні шари II і IV складаються в основному із сприймаючих елементів і є «входами» в кору. Крупноклеточний шар V — вихід з кори в підкірку, а среднеклеточний шар III — асоціативний, зв'язуючий між собою різні кіркові зони ( див. мал.(малюнок) 1 ) .
Локалізація функцій в корі характеризується динамічністю внаслідок того, що, з одного боку, є строго локалізовані і просторово відмежовані зони кора, пов'язана із сприйняттям інформації від певного органу чуття, а з іншої — кора є єдиним апаратом, в якому окремі структури тісно зв'язані і у разі потреби можуть взаємозамінюватися (т.з. пластичність кіркових функцій). Крім того, в кожен даний момент кіркові структури (нейрони, поля, області) можуть утворювати комплекси, що погоджено діють, склад яких змінюється залежно від специфічних і неспецифічних стимул-реакцій, що визначають розподіл гальмування і збудження в корі. Нарешті, існує тісна взаємозалежність між функціональним станом кіркових зон і діяльністю підкіркових структур. Території кори різко розрізняються по своїх функціях. Велика частина древньої кори входить в систему нюхового аналізатора. Стара і проміжна кора, будучи тісно пов'язаними з древньою корою як системами зв'язків, так і еволюційно, не мають прямого відношення до нюху. Вони входять до складу системи, регуляцією вегетативних реакцій і емоційних станів організму, що відає (див. Ретикулярна формація, Лімбічеськая система ) . Нова кора — сукупність кінцевих ланок різних сприймаючих (сенсорних) систем (кіркових кінців аналізаторів ) .
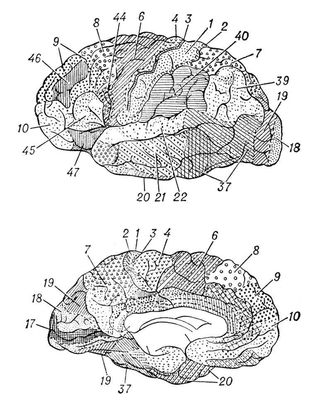
Прийнято виділяти в зоні того або іншого аналізатора проекційні, або первинні, і вторинні, поля, а також третинні поля, або асоціативні зони. Первинні поля отримують інформацію, опосередковану через найменшу кількість перемикань в підкірці (у зоровому горбі, або таламусі, проміжного мозку). На цих полях як би спроектована поверхня периферичних рецепторів ( мал. 4 ) . В світлі сучасних даних, проекційні зони не можна розглядати як пристрої, що сприймають роздратування «крапку в крапку». У цих зонах відбувається сприйняття певних параметрів об'єктів, тобто створюються (інтегруються) образи, оскільки дані ділянки мозку відповідають на певні зміни об'єктів, на їх форму, орієнтацію, швидкість руху і т. п.
Крім того, локалізація функцій в первинних зонах багато разів дублюється по механізму, що нагадує голографію, коли кожна найменша ділянка пристрою, що запам'ятовує містить відомості про весь об'єкт. Тому досить збереження невеликої ділянки первинного сенсорного поля, щоб здібність до сприйняття майже повністю збереглася. Вторинні поля отримують проекції від органів чуття через додаткові перемикання в підкірці, що дозволяє виробляти складніший аналіз того або іншого образу. Нарешті, третинні поля, або асоціативні зони, отримують інформацію від неспецифічних підкіркових ядер, в яких підсумовується інформація від декількох органів чуття, що дозволяє аналізувати і інтегрувати той або інший об'єкт в ще абстрагованішій і узагальненій формі. Ці області називаються також зонами перекриття аналізаторів. Первинні і частково вторинні поля — можливий субстрат першої сигнальної системи, а третинні зони (асоціативні) — другою сигнальною системи, специфічною для людини (І. П. Павлов). Ці структури міжаналізаторів визначають складні форми мозкової діяльності, включаючі і професійні навики (ніжнетеменная область), і мислення, планерування і цілеспрямованість дій (лобова область), і письмову і усну мову (нижня лобова підобласть, скронева, скронево-тім'яно-потилична і ніжнетеменная області). Основні представники первинних зон в потиличної області — поле 17, де спроектована сітківка, в скроневій — поле 41, де спроектований Кортієв орган, в прецентральной області — поле 4, де здійснюється проекція пропріорецепторів відповідно до розташування мускулатури, в центральній для поста — поля 3 і 1, де спроектовані екстерорецепції відповідно до їх розподілу в шкірі. Вторинні зони представлені полями 8 і 6 (руховий аналізатор ) , 5 і 7 (шкірний аналізатор), 18 і 19 (зоровий аналізатор), 22 (слуховий аналізатор ). Третинні зони представлені обширними ділянками лобової області (поля 9, 10, 45, 44 і 46), ніжнетеменной (поля 40 і 39), скронево-тім'яно-потиличною (поле 37).
Кіркові структури грають первинну роль у вченні тварин і людини. Проте освіта деяких простих умовних рефлексів, головним чином з внутрішніх органів, може бути забезпечено підкірковими механізмами. Ці рефлекси можуть утворюватися і на нижчих рівнях розвитку, коли ще немає кори. Складні умовні рефлекси, лежачі в основі цілісних актів поведінка, вимагають збереження кіркових структур і участі не лише первинних зон кіркових кінців аналізаторів, але і асоціативних — третинних зон. Кіркові структури мають пряме відношення і до механізмів пам'яті . Електророздратування окремих областей кори (наприклад, скроневою) викликає у людей складні картини спогадів.
Характерна особливість діяльності кори — її спонтанна електрична активність, що реєструється у вигляді електроенцефалограми (ЕЕГ). В цілому кора і її нейрони володіють ритмічною активністю, яка відображає ті, що відбуваються в них біохімічні і біофізичні процеси. Ця активність має всіляку амплітуду і частоту (від 1 до 60 гц ) і змінюється під впливом різних чинників.
Ритмічна активність кори нерегулярна, проте можна по частоті потенціалів виділити декілька різних типів її (альфа-, бета-, дельта- і тета-рітмі). ЕЕГ зазнає характерні зміни при багатьох фізіологічних і патологічних станах (різних фазах сну, при пухлинах, судорожних припадках і т. і.). Ритм, тобто частота, і амплітуда біоелектричних потенціалів кори задаються підкірковими структурами, які синхронізують роботу груп кіркових нейронів, що і створює умови для їх погоджених розрядів. Цей ритм пов'язаний з апікальнимі (верхівковими) дендритом пірамідних кліток. На ритмічну діяльність кори накладаються впливи, що йдуть від органів чуття. Так, спалах світла, клацання або дотик до шкіри викликають у відповідних зонах т.з. первинна відповідь, що складається з ряду позитивних хвиль (відхилення електронного променя на екрані осцилографа вниз) і негативної хвилі (відхилення світивши вгору). Ці хвилі відображають діяльність структур даної ділянки кори і міняються в її різних шарах.
Філогенез і онтогенез кори. Кора — продукт тривалого еволюційного розвитку, в процесі якого спочатку з'являється древня кора, що виникає у зв'язку з розвитком нюхового аналізатора у риб. З виходом тварин з води на сушу починає інтенсивно розвиватися т.з. плащевідная, повністю відособлена від підкірки частина кори, яка складається із старої і нової кори. Становлення цих структур в процесі пристосування до складних і всіляких умов наземного існування зв'язано (вдосконаленням і взаємодією різних сприймаючих і рухових систем. В земноводних кора представлена древньою і зачатком старої кори, у плазунів добре розвинені древня і стара кора і з'являється зачаток нової кори. Найбільшого розвитку нова кора досягає у ссавців, а серед них в приматів (мавпи і людина), хоботних (слони) і китоподібних (дельфіни, кити). У зв'язку з нерівномірністю зростання окремих структур нової кори її поверхня стає складчастою, покриваючись борознами і звивиною. Вдосконалення кори кінцевого мозку у ссавців нерозривно пов'язане з еволюцією всіх відділів центральної нервової системи. Цей процес супроводиться інтенсивним зростанням прямих і зворотних зв'язків, що сполучають кіркові і підкіркові структури. Т. о., на вищих етапах еволюції функції підкіркових утворень починають контролюватися кірковими структурами. Дане явище отримала назва кортіколізациі функцій. В результаті кортіколізациі ствол мозку утворює з кірковими структурами єдиний комплекс, а пошкодження кори на вищих етапах еволюції приводить до порушення життєво важливих функцій організму. Найбільші зміни і збільшення в процесі еволюції нової кори зазнають асоціативні зони, тоді як первинні, сенсорні поля зменшуються по відносній величині. Розростання нової кори приводить до витіснення старою і древньою на нижню і серединну поверхні мозку.
Кіркова пластинка з'являється в процесі внутріутробного розвитку людини порівняно рано — на 2-м-коді місяці. Раніше всього виділяються нижні шари кори (VI—VII), потім — вище розташовані (V, IV, III і II; див. мал.(малюнок) 1 ). До 6 місяців у ембріона вже є всі цитоархитектонічеськие поля кори властиві дорослій людині. Після народження в зростанні кори можна виділити три переломні етапи: на 2—3-м-коді місяці життя, в 2,5—3 роки і в 7 років. До останнього терміну цитоархитектоника кори повністю сформована, хоча тіла нейронів продовжують збільшуватися до 18 років. Кіркові зони аналізаторів завершують свій розвиток раніше, і міра їх збільшення менша, ніж у вторинних і третинних зон. Наголошується велика різноманітність в термінах дозрівання кіркових структур в різних індивідуумів, що збігається з різноманітністю термінів дозрівання функціональних особливостей кори. Т. о., індивідуальне (онтогенез ) і історичне (філогенез ) розвиток кори характеризується схожими закономірностями.
Літ.: Орбелі Л. А., Питання вищої нервової діяльності, М.— Л., 1949; Цитоархитектоника кори великого мозку людини. Сб. ст., М., 1949; Філімонов І. Н., Порівняльна анатомія кори великого мозку ссавців, М., 1949; Павлов І. П., Двадцятирічний досвід об'єктивного вивчення вищої нервової діяльності тварин, Полн. собр. соч.(вигадування), 2 видавництва, т. 3, кн. 1—2, М., 1951; Брейзье М., Електрична активність нервової системи, пер.(переведення) з англ.(англійський), М., 1955; Сепп Е. До., Історія розвитку нервової системи хребетних, 2 видавництва, М., 1959; Лурія А. Р., Вищі кіркові функції людини і їх порушення при локальних ураженнях мозку, М., 1962; Воронін Л. Р., Курс лекцій з фізіології вищої нервової діяльності, М., 1965; Поляків Р. І., Про принципи нейронної організації мозку, М., 1965; Кіркова регуляція діяльності підкіркових утворень головного мозку. Сб. ст., Тб., 1968; Анохин П. До., Біологія і нейрофізіологія умовного рефлексу, М., 1968; Берітов І. С., Структура і функції кори великого мозку, М., 1969.