Ретикулярна формація (від латів.(латинський) reticulum — сіточка, formatio — освіта), сітьовидна освіта, сукупність нервових структур, розташованих в центральних відділах стволової частини мозку (довгастому і середньому мозку, зорових горбах). Нейрони, складові Р. ф., всілякі по величині, будові і довжині аксонів ; їх волокна густо переплітаються. Термін «Р. ф.», введений німецьким вченим О. Дейтерсом, відображає лише морфологічні її особливості. Р. ф. морфологічно і функціонально пов'язана із спинним мозком, мозочком, лімбічеськой системою і корою великих півкуль головного мозку. В області Р. ф. здійснюється взаємодія що поступають в неї як висхідних — аферентних, так і низхідних — еферентних імпульсів. Можлива також циркуляція імпульсів по замкнутих нейронних ланцюгах. Т. о., існує постійний рівень збудження нейронів Р. ф., унаслідок чого забезпечуються тонус і певна міра готовності до діяльності різних відділів центральної нервової системи. Міра збудження Р. ф. регулюється корою великих півкуль головного мозку .
Низхідні впливи. У Р. ф. розрізняють області, які роблять гальмівні і полегшуючі впливи на рухові реакції спинного мозку ( мал. 1 ). Залежність між роздратуванням різних областей ствола мозку і спинномозковими рефлексами вперше відзначив в 1862 І. М. Сеченов . В 1944—46 американський нейрофізіолог Х. Мегоун із співробітниками показали, що роздратування різних ділянок Р. ф. довгастого мозку робить полегшуючий або гальмівний вплив на рухові реакції спинного мозку. Електричне роздратування медіальної частини Р. ф. довгастого мозку в наркотизованих і децеребрірованних кішок і мавп супроводиться повним припиненням рухів, що викликаються як рефлекторно, так і стимуляцією рухових ділянок кори мозку. Всі гальмівні ефекти — двосторонні, але на стороні роздратування такий ефект незрідка спостерігається при нижчому порозі роздратування. Деякі прояви гальмівних впливів Р. ф. довгастого мозку відповідають картині центрального гальмування, описаного Сеченовим (див. Сеченовськоє гальмування ) . Роздратування латеральної області Р. ф. довгастого мозку по периферії області, що робить гальмівні впливи, супроводиться полегшуючою дією на моторну активність спинного мозку. Область Р. ф., що робить полегшуючі впливи на спинний мозок, не обмежується довгастим мозком, а поширюється наперед, захоплюючи область варолієва моста і середнього мозку. Р. ф. може впливати на різні утворення спинного мозку, наприклад на альфа-мотонейрони, іннервуючі основні (екстрафузальниє) волокна м'язів, що беруть участь в довільних рухах. Збільшення латентних періодів відповідей мотонейронів при роздратуванні гальмівних відділів Р. ф. дозволяє передбачати, що гальмівні впливи ретикулярних структур на рухові реакції спинного мозку здійснюються за допомогою вставних нейронів, можливо кліток Реншоу. Механізм впливу Р. ф. на м'язовий тонус розкритий шведським нейрофізіологом Р. Гранітом що показав, що Р. ф. впливає також на активність гамма-мотонейронів, аксони яких йдуть до так званих інтрафузальним м'язовим волокнам, здійснюючи важливу роль в регуляції пози і фазних рухів організму.
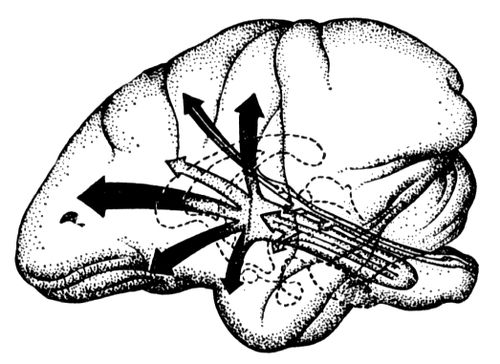
Висхідні впливи. Різні відділи Р. ф. (від проміжного до довгастого мозку) роблять збуджуючі генералізованниє впливи на кору мозку, тобто залучають до процесу збудження всі області кори великих півкуль ( мал. 2 ) . В 1949 італійський фізіолог Дж. Моруцци і Мегоун, досліджуючи біоелектричну активність мозку, встановили, що роздратування Р. ф. ствола мозку змінює повільні синхронні високовольтні коливання, характерні для сну, на нізкоамплітудную високочастотну активність, характерну для неспання. Зміна електричній активності кори мозку супроводиться у тварин зовнішніми проявами пробудження. Р. ф. тісно пов'язана анатомічно з класичними провідними дорогами, і збудження її здійснюється за допомогою екстеро- і інтероцептивних аферентних (чутливих) систем. На цій підставі ряд авторів відносить Р. ф. до неспецифічної аферентної системи мозку. Проте вживання різних фармакологічних речовин при вивченні функції Р. ф., відкриття виборчої дії хімічних препаратів на реакції, здійснювані за участю Р. ф., дозволили П. До. Анохину сформулювати положення про специфічність висхідних впливів Р. ф. на кору мозку. Активуючі впливи Р. ф. завжди мають певне біологічне значення і характеризуються виборчою чутливістю до різних фармакологічних речовин (Анохин, 1959, 1968). Введені в організм наркотичні засоби викликають гальмування нейронів Р. ф., блокуючи тим самим її висхідні активуючі впливи на кору мозку.
Важлива роль в підтримці активності Р. ф., чутливих до різних циркулюючих в крові хімічних речовин, належить гуморальним чинникам: катехоламінам, двоокису вуглецю, холінергичеським речовинам і т. д. Це забезпечує включення Р. ф. у регулювання деяких вегетативних функцій. Кора великих півкуль, що випробовує тонічні активуючі впливи з боку Р. ф., може активно змінювати функціональний стан ретикулярних утворень (змінювати швидкість проведення збудження в ній, впливати на функціонування окремих нейронів), тобто контролювати, по вираженню І. П. Павлова, «сліпу силу» підкірки.
Відкриття властивостей Р. ф., її взаємин з іншими підкірковими структурами і областями кори мозку дозволило уточнити нейрофізіологічні механізми болю, сну, неспання, активної уваги, формування цілісних умовно-рефлекторних реакцій, розвитку різних мотиваційних і емоційних станів організму. Дослідження Р. ф. з використанням фармакологічних засобів відкривають можливості медикаментозного лікування ряду захворювань центральної нервової системи, обумовлюють новий підхід до таких найважливіших проблем медицини, як наркоз і ін.
Літ.: Бродал А., Ретикулярна формація мозкового ствола, пер, з англ.(англійський), М., 1960; Россі Дж. Ф., Цанкетті А., Ретикулярна формація ствола мозку, пер.(переведення) з англ.(англійський) М., 1960; Ретикулярна формація мозку, пер.(переведення) з англ.(англійський), М., 1962; Мегун Р., Безсонний мозок, пер.(переведення) з англ.(англійський), 2 видавництва, М., 1965; Анохин П. До., Біологія і нейрофізіологія умовного рефлексу, М., 1968; Гранує Р., Основи регуляції рухів, пер.(переведення) з англ.(англійський), М., 1973; Moruzzi G., Magoun Н. W., Brain stem reticular formation and activation of EEG, в книга: Electroencephalography and clinical neurophysiology, v. 1, Boston 1949