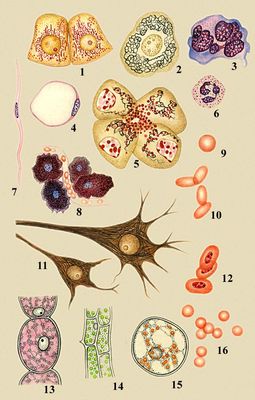
Клітка, елементарна жива система, здібна до самостійного існування, самовідтворення і розвитку; основа будови і життєдіяльності всіх тварин і рослин. До. існують і як самостійні організми> (див. Прості ), і у складі багатоклітинних організмів (тканинні До.). Термін «До.» запропонований англійським мікроськопістом Р. Гуком (1665). До. — предмет вивчення особливого розділу біології — цитології . Систематичне вивчення До. почалося лише в 19 ст Одним з найбільших наукових узагальнень того часу була клітинна теорія, що затверджувала єдність будови всієї живої природи. Вивчення життя на клітинному рівні лежить в основі сучасних біологічних досліджень.
В будові і функціях кожній До. виявляються ознаки, загальні для всіх До., що відображає єдність їх походження з первинних органічних комплексів. Приватні особливості різних До. — результат їх спеціалізації в процесі еволюції. Так, все До. схоже регулюють обмін речовин, подвоюють і використовують свій спадковий матеріал, отримують і утилізували енергію. В той же час різні одноклітинні організми (амеби, інфузорії і т.д.) сильно розрізняються розмірами, формою поведінкою. Не менше різкий розрізняються До. багатоклітинних організмів. Так, у людини є лімфоїдні До. — невеликі (діаметром близько 10 мкм ) округлі До., що беруть участь в імунологічних реакціях, і нервові До., частина яких має відростки завдовжки більше метра; ці До. здійснюють основні регуляторні функції в організмі.
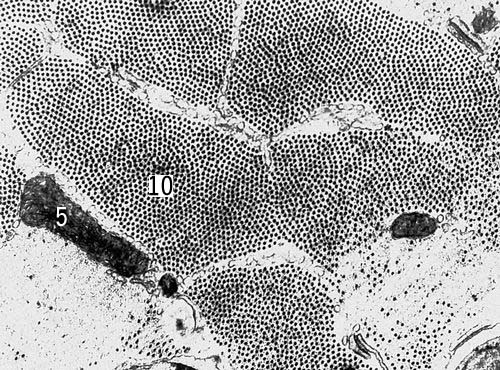
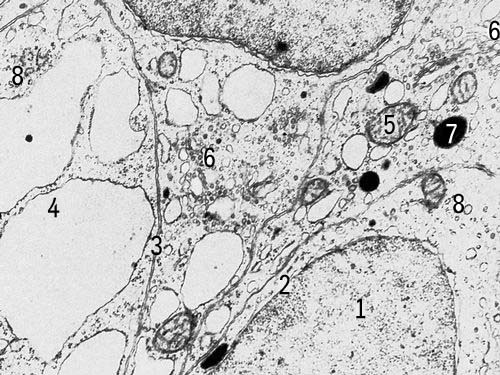
Методи дослідження. Першим цитологичеським методом була мікроскопія живих До. Сучасні варіанти прижиттєвої (вітальною) світлової мікроскопії — фазово-контрастна, люмінесцентна, інтерференційна і ін. (див. Мікроскоп ) — дозволяють вивчати форму. і загальна будова деяких її структур, рух До. і їх ділення. Деталі будови До. виявляються лише після спеціального контрастування, що досягається забарвленням убитої К. Новий етап вивчення структури До. — електронна мікроскопія, що дає значно більший дозвіл структур До. в порівнянні зі світловою мікроскопією (див. Роздільна здатність оптичних приладів). Хімічний склад До. вивчається цито- і гістохімічними методами, що дозволяють з'ясувати локалізацію і концентрацію речовин в клітинних структурах, інтенсивність синтезу речовин і їх переміщення в До. (див. Гистохимія ). Цитофізіологичеськие методи дозволяють вивчати функції., наприклад збудження, секрецію. Див. також Авторадіографія,Мікроскопічна техніка,Цитофотометрія .
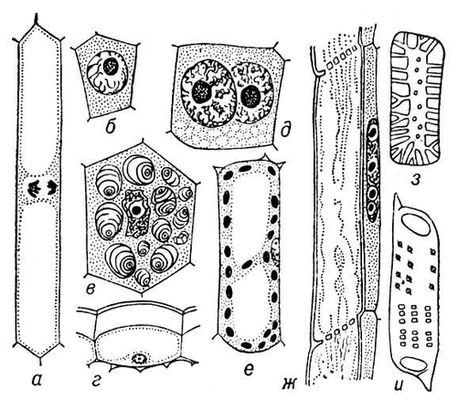
Загальні властивості кліток. В кожній До. розрізняють дві основні частини — ядро і цитоплазму, в яких, у свою чергу, можна виділити структури, що розрізняються формою, розмірам, внутрішній будові, хімічним властивостям і функціям. Одні з них — так звані органоїди — життєво необхідні До. і виявляються у всіх К. Другие — продукти активності До., представляючі тимчасові утворення. У спеціалізованих структурах здійснюється розділення різних біохімічних функцій, що сприяє здійсненню в одній і тій же До. різнорідних процесів, включаючий синтез і розпад багатьох речовин.
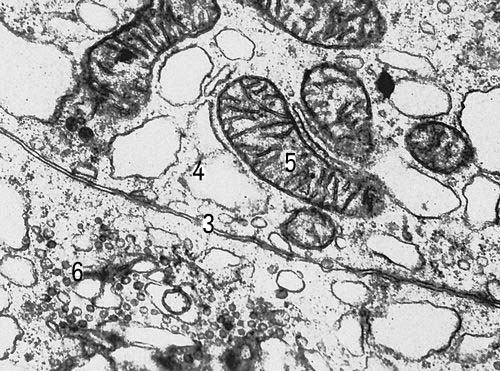
В ядерних органоїдах — хромосомах, в їх основному компоненті — ДНК(дезоксирибонуклеїнова кислота), зберігається генетична інформація про будову білків, властивих організму певного вигляду (див. Ген,Генетичний код ). Інша найважливіша властивість ДНК(дезоксирибонуклеїнова кислота) — здібність до самовідтворення, що забезпечує як стабільність спадкової інформації, так і її безперервність — передачу наступним поколінням. На обмежених ділянках ДНК(дезоксирибонуклеїнова кислота), що охоплює декілька генів, як на матрицях, синтезуються рибонуклеїнові кислоти — безпосередні учасники синтезу білка. Перенос (транскрипція ) коди ДНК(дезоксирибонуклеїнова кислота) відбувається при синтезі інформаційних РНК(рибонуклеїнова кислота) (І-РНК). Синтез білка представляється як прочитування інформації з матриці РНК(рібонуклеїнова кислота). У цьому процесі, званому трансляцією, беруть участь транспортні РНК(рибонуклеїнова кислота) (Т-РНК) і спеціальні органоїди — рибосоми, що утворюються в ядерці . Розміри ядерця визначаються головним чином потребою До. у рибосомах; тому особливо велике воно в До., інтенсивно синтезуючих білок. Синтез білка — кінцевий висновок реалізації функцій хромосом — здійснюється головним чином в цитоплазмі. Білки — ферменти, деталі структур і регулювальники різних процесів, включаючи і транскрипцію — визначають кінець кінцем всі сторони життя До., дозволяючи До. зберігати свою індивідуальність, не дивлячись на постійно змінне оточення. Якщо в бактерійній До. синтезується близько 1000 різних білків, то майже в кожній з До. людини — зверху 10000. Таким чином, різноманітність внутріклітинних процесів в ході еволюції організмів істотно зростає. Оболонка ядра, що відокремлює його вміст від цитоплазми, складається з двох мембран, пронизаних порами, — спеціалізованих ділянок для транспорту деяких з'єднань з ядра в цитоплазму і назад. Інші речовини проходять через мембрани шляхом дифузії або активного транспорту, що вимагає витрат енергії. Багато процесів відбуваються в цитоплазмі До. при участі мембран ендоплазматичній мережі — основної синтезуючої системи До., а також Гольджі комплексу і мітохондрій . Відмінності мембран різних органоїдів визначаються властивостями створюючих їх білків і ліпідів. До деяких мембран ендоплазматичної мережі прикріплені рибосоми; тут відбувається інтенсивний синтез білка. Така гранулярна ендоплазматична мережа особливо розвинена в До., що секретують або інтенсивно оновлюючих білок, наприклад у людини в До. печінки, підшлункової залози, нервових До. До складу інших біологічних мембран, позбавлених рибосом (гладкоконтурная мережа), входять ферменти, що беруть участь в синтезі вуглеводно-білкових і ліпідних комплексів. У каналах ендоплазматичній мережі можуть тимчасово накопичуватися продукти діяльності К.; в деяких До. по каналах відбувається направлений транспорт речовин. Перед виведенням з До. речовини концентруються в пластинчастому комплексі (комплексі Гольджі). Тут відособляються різні включення До., наприклад секреторні або пігментні гранули, утворюються лізосоми — бульбашки, що містять гідролітичні ферменти і що беруть участь в внутріклітинному переварюванні багатьох речовин. Система оточених мембранами каналів, вакуолей і бульбашок представляє одне ціле. Так, ендоплазматична мережа може без перерви переходити в мембрани, що оточують ядро, з'єднуватися з цитоплазматичною мембраною, формувати комплекс Гольджі. Проте зв'язки ці нестабільні. Незрідка, а в багатьох До. зазвичай різні мембранні структури роз'єднані і обмінюються речовинами через гіалоплазму . Енергетика До. багато в чому залежить від роботи мітохондрій. Число їх вагається в До. різного типа від десятків до тисяч. Наприклад, в печінковій До. людини близько 2 тис. мітохондрій; їх загальний об'єм не менше 1 / 5 об'єму К. Внешняя мембрана мітохондрії відмежовує її від цитоплазми, на внутрішній — відбуваються основні енергетичні перетворення речовин, в результаті яких утворюється з'єднання, багате енергією, — аденозинтрифосфорна кислота (АТФ) — універсальний переносник енергії в К. Мітохондрії містять ДНК(дезоксирибонуклеїнова кислота) і здібні до самовідтворення; проте автономність мітохондрій відносна, їх репродукція і діяльність залежать від ядра. За рахунок енергії АТФ в До. здійснюються різні синтези, транспорт і виділення речовин, механічна робота, регуляція процесів і т.д. У діленні До. і інколи в їх русі беруть участь структури, що мають вигляд трубочок субмікроскопічних розмірів. «Збірка» таких структур і їх функціонування залежать від центріолей, за участю яких організовується веретено ділення клітки, з чим пов'язано переміщення хромосом і орієнтацію осі ділення К. Базальниє тільця — похідні центріолей — необхідні для побудови і нормальної роботи джгутиків і вій — локомоторних і чутливих утворень До., будова яких в простих і в різних До. багатоклітинних однотипно.
Від позаклітинного середовища До. відокремлена плазматичною мембраною, через яку відбувається вступ іонів і молекул в До. і виділення їх з К. Отношеніє поверхні До. до її об'єму зменшується із збільшенням об'єму, і чим більше До., тим більше утруднені її зв'язки із зовнішнім середовищем. Величина До. не може бути особливе великою. Для живих До. характерний активний транспорт іонів, що вимагає витрати енергії, спеціальних ферментів і, можливо, переносників. Завдяки активному і виборчому перенесенню в До. одних іонів і безперервному видаленню з неї інших створюється різниця концентрацій іонів в До. і довкіллю. Цей ефект може бути обумовлений і скріпленням іонів компонентамі К. Многие іони необхідні як активатори внутріклітинних синтезів і як стабілізатори структури органоїдів. Оборотні зміни співвідношення іонів в До. і середовищу лежать в основі біоелектричної активності До. — одного з важливих чинників передачі сигналів від однієї До. до іншої (см.Біоелектричні потенціали ). Утворюючи впячиванія, які потім замикаються і відділяються у вигляді бульбашок всередину До., плазматична мембрана здатна захоплювати розчини великих молекул (піноцитоз ) або навіть окремі частки величиною в декілька мкм (фагоцитоз ). Так здійснюється живлення деяких До., перенесення речовин через До., захват бактерій фагоцитами. З властивостями плазматичної мембрани зв'язані і сили зчеплення, що утримують у багатьох випадках До. один біля одного, наприклад в покривах тіла або внутрішніх органах. Зчеплення і зв'язок До. забезпечуються хімічною взаємодією мембран і спеціальними структурами мембрани — десмосомами .
Розглянута в загальній формі схема будови До. властива в основних межах як тваринам, так і рослинним До.. Але є і істотні відмінності в особливостях метаболізму і будови рослинних До. від тварин.
Клітки рослин . Поверх плазматичної мембрани рослинні До. покриті, як правило, твердою зовнішньою оболонкою (вона може бути відсутньою лише в статевих До.), що полягає у більшості рослин головним чином з полісахаридів: целюлози, пектинових речовин і геміцеллюлоз, а в грибів і деяких водоростей — з хітину. Оболонки забезпечені порами, через які за допомогою виростов цитоплазми сусідні До. зв'язані один з одним. Склад і будова оболонки міняються по мірі зростання і розвитку К. Часто в До., що припинили зростання, оболонка просочується лігніном, кремнеземом або ін. речовиною, яка робить її міцнішою. Оболонки До. визначають механічні властивості рослини. До. деяких рослинних тканин відрізняються особливо товстими і міцними стінками (див. Деревина ), що зберігають свої скелетні функції після загибелі К. Діфференцированниє рослинні До. мають декілька вакуолею або одну центральну вакуоль, що займає зазвичай велику частину об'єму К. Содержімоє вакуолею, — розчин різних солей, вуглеводів, органічних кислот, алкалоїдів, амінокислот, білків, а також запас води. У вакуолях можуть відкладатися живильні речовини. У цитоплазмі рослинної До. є спеціальні органоїди — пластиди ; лейкопласт (у них часто відкладається крохмаль), хлоропласти (містять переважно хлорофіл і здійснюють фотосинтез ) і хромопласти (містять пігменти з групи каротиноїдів). Пластиди, як і мітохондрії, здібні до самовідтворення. Комплекс Гольджі в рослинній До. представлений розсіяними по цитоплазмі діктіосомамі .
Одноклітинні організми. В будові і функціях одноклітинних, або простих, межі, властиві будь-який До., поєднуються з ознаками самостійних організмів. Так, в простих такий же набір органоїдів, як і в До. багатоклітинних; ідентично і ультрастроєніє їх органоїдів; при діленні простих в них виявляються типові хромосоми. Проте пристосування простих до різних місць існування (водною або наземною, до вільного або паразитичному існуванню) зумовило істотну різноманітність їх будови і фізіології. Багато простих (жгутикові, інфузорії) володіють складним руховим апаратом і мають органели, пов'язані із захватом їжі і травленням. Вивчення простих представляє великий інтерес для з'ясування можливостей філогенезу К.: еволюційні зміни організму протікають у них на клітинному рівні. На відміну від простих і К. багатоклітинних організмів, бактерії синезелениє водорості, актиноміцети не мають оформленого ядра і хромосом. Їх генетичний апарат, називається нуклеоїдом, представлений нитками ДНК(дезоксирибонуклеїнова кислота) і не оточений оболонкою. Ще більш відрізняються від До. багатоклітинних організмів і від простих віруси, в яких відсутні основні, необхідні для обміну речовин ферменти. Тому віруси можуть зростати і розмножуватися, лише проникаючи в До. і використовуючи їх ферментні системи.
Спеціальні функції кліток. В процесі еволюції багатоклітинних виникло розділення функцій між До., що привело до розширення можливостей пристосування тварин і рослин до змінних умов середовища. Що закріпилися спадково відмінності у формі До., їх розмірах і деяких сторонах метаболізму реалізуються в процесі індивідуального розвитку організму. Основний прояв розвитку — диференціювання До., їх структурна і функціональна спеціалізація. Диференційовані До. мають такий же набір хромосом, як і запліднена яйцеклітина. Це доводиться пересадкою ядра диференційованої До. у заздалегідь позбавлену ядра яйцеклітину, після чого може розвиватися повноцінний організм. Таким чином, відмінності між диференційованими До., мабуть, обумовлюються різними співвідношеннями активних і неактивних генів, кожен з яких кодує біосинтез певного білка. Судячи по складу білків, в диференційованих До. активна (здібна до транскрипції) лише невелика частина (порядка 10%) генів, властивих До. даного вигляду організмів. Серед них лише небагато відповідальні за спеціальну функцію До., а останні забезпечують загальноклітинні функції. Так, в мишечних До. активні гени, що кодують структуру скоротних білків, в ерітроїдних До. — гени що кодують біосинтез гемоглобіну, і т.д. Проте в кожній До. мають бути активні гени, що визначають біосинтез речовин і структур, необхідних для всіх До., наприклад ферментів, що беруть участь в енергетичних перетвореннях речовин. В процесі спеціалізації До. окремі загальноклітинні функції їх можуть розвиватися особливо сильно. Так, в залізистих До. понад усе виражена синтетична активність, мишечниє — найбільш сократіми, нервові, — найбільш збудливі. У вузькоспеціалізованих До. виявляються структури, характерні лише для цих До. (наприклад, у тварин — міофібрили м'язів, тонофібрили і вії деяких покривних До., нейрофібрили нервових До., джгутики в простих або в сперматозоїдів багатоклітинних організмів). Інколи спеціалізація супроводиться втратою деяких властивостей (наприклад, нервові До. втрачають здібність до розмноження; ядра До. кишкового епітелію ссавців не можуть в зрілому стані синтезувати РНК(рибонуклеїнова кислота); зрілі еритроцити ссавців позбавлені ядра). Виконання важливих для організму функцій включає інколи загибель До. Так, До. епідермісу шкіри поступово ороговевают і гинуть, але залишаються деякий час в пласті, оберігаючи ті, що підлягають тканині від пошкодження і інфекції. У сальних залозах До. поступово перетворюються на краплі жиру, який використовується організмом або виділяється. Для виконання деяких тканинних функцій До. утворюють неклітинні структури. Основні дороги їх освіти — секреція або перетворення компонентів цитоплазми. Так, значна за об'ємом частина підшкірної клітковини, хряща і кісті складає проміжну речовину — похідне До. сполучній тканині. До. крові мешкають в рідкому середовищі (плазмі крові), що містить білки, цукру і ін. речовини, що виробляються різними До. організму. До. епітелію, створюючі пласт, оточені тонким прошарком дифузно розподілених речовин, головним чином глікопротеїдів (так званий цемент, або надмембранний компонент). Зовнішні покриви членистоногих і раковини молюсків — також продукти виділення К. Взаїмодействіє спеціалізованих До. — необхідна умова життя організму і незрідка самих цих До. (див. Гістологія ) . Позбавлені зв'язків один з одним, наприклад в культурі, До. швидко втрачають особливості властивих їм спеціальних функцій.
Ділення клітин . У основі здатності До. до самовідтворення лежать унікальна властивість ДНК(дезоксирибонуклеїнова кислота) самокопіроваться і строге рівноцінне ділення репродукованих хромосом в процесі мітоза . В результаті ділення утворюються дві До., ідентичні початковою по генетичних властивостях і з оновленим складом ядра і цитоплазми. Процеси самовідтворення хромосом, їх ділення, утворення двох ядер і ділення цитоплазми розділені в часі, складаючи в сукупності мітотичний цикл До. У випадку, якщо після ділення До. починає готуватися до наступного ділення, мітотичний цикл збігається з життєвим циклом К. Однако у багатьох випадках після ділення (а інколи перед ним) До. виходять з мітотичного циклу, диференціюються і виконують в організмі ту або іншу спеціальну функцію. Склад таких До. може оновлюватися за рахунок ділень малодиференційованих До. У деяких тканинах і диференційовані До. здатні повторно входити в мітотичний цикл. У нервовій тканині диференційовані До. не діляться; багато хто з них живе так само довго, як організм в цілому, тобто у людини — декілька десятків років. При цьому ядра нервових До. не втрачають здібності до ділення: будучи пересаджені в цитоплазму ракових До., ядра нейронів синтезують ДНК(дезоксирибонуклеїнова кислота) і діляться. Досліди з клітками-гібридами показують вплив цитоплазми на прояв ядерних функцій. Неповноцінна підготовка до ділення запобігає мітоз або спотворює його течію. Так, в деяких випадках не відбувається ділення цитоплазми і утворюється двуядерная К. Многократноє ділення ядер в тій, що не ділиться До. приводить до появи багатоядерних До. або складних надклеточних структур (симпластов), наприклад в поперечнополосатих м'язах. Інколи репродукція До. обмежується відтворенням хромосом, і утворюється поліплоїдная До., що має подвоєний (порівняно до початкової До.) набір хромосом. Поліплоїдізация приводить до посилення синтетичної активності, збільшенню розмірів і маси До.
Оновлення кліток. Для тривалої роботи кожній До. необхідне відновлення зношуваних структур, як і ліквідація пошкоджень До., викликаних зовнішніми діями. Відновні процеси, характерні для всіх До., пов'язані із змінами проникності плазматичної мембрани і супроводяться посиленням внутріклітинних синтезів, в першу чергу синтезу білка. У багатьох тканинах стимуляція відновних процесів приводить до репродукції генетичного апарату і ділення К.; це властиво, наприклад покривам або кровотворній системі. Процеси внутріклітинного оновлення в цих тканинах виражені слабо, їх До. живуть порівняно недовго (наприклад, До. кишкового покриву ссавців — всього декілька діб). Максимальної вираженості внутріклітинні відновні процеси досягають в тих, що не діляться або слабоделящихся клітинних популяціях, наприклад в нервових К. Показателем досконалість процесів внутрішнього оновлення До. є тривалість їх життя; для багатьох нервових До. вона збігається з тривалістю життя всього організму.
Мутації. Зазвичай процес відтворення ДНК(дезоксирибонуклеїнова кислота) відбувається без відхилень, і генетичний код залишається постійним, що забезпечує синтез одного і того ж набору білків у величезному числі клітинних поколінь. Проте в окремих випадках може статися мутація — часткова зміна структури гена. Кінцевий її ефект — зміна властивостей білків, що кодуються генами мутантів. Якщо при цьому зачіпаються важливі ферментні системи, властивості До., а інколи і всього організму істотно змінюються. Так, мутація одного з генів, контролюючих синтез гемоглобіну, приводить до важкого захворювання — анемії . Природний відбір корисних мутацій — важливий механізм еволюції.
Регуляція функцій кліток. Основний механізм регуляції внутріклітинних процесів пов'язаний з різними впливами на ферменти — високо специфічні каталізатори біохімічних реакцій. Регуляція може здійснюватися на генетичному рівні, коли визначається склад ферментів або кількість того або іншого ферменту в До. У останньому випадку регуляція може відбуватися і на рівні трансляції. Інший тип регуляції — дія на сам фермент, внаслідок чого може відбуватися як гальмування, так і стимуляція його активності. Структурний рівень регуляції — вплив на збірку клітинних структур: мембран, рибосом і т.д. Конкретними регулювальниками внутріклітинних процесів можуть бути нервові впливи, гормони, спеціальні речовини, що виробляються усередині До. або що оточують До. (особливо білки), або ж самі продукти реакцій. У останньому випадку дія здійснюється за принципом зворотного зв'язку, коли продукт реакції впливає на активність ферменту — каталізатора цієї реакції. Регуляція може здійснюватися через транспорт попередників і іонів, впливу на матричний синтез (РНК, полісоми, ферменти синтезу), зміну форми регульованого ферменту.
Організація і регуляція функцій До. на молекулярному рівні визначають такі властивості живих систем, як просторова компактність і енергетична економічність. Важлива властивість багатоклітинних організмів — надійність — багато в чому залежить від множинності (взаємозамінюваності) До. кожного функціонального типа, а також від можливості їх заміни в результаті розмноження До. і оновлення компонентів кожній До.
В медицині використовуються дії на До. для лікування і запобігання захворюванням. Багато лікарських речовин змінюють активність визначених До. Так, наркотики, транквілізатори і болезаспокійливі речовини знижують інтенсивність діяльності нервових До., а стимулятори її підсилюють. Деякі речовини стимулюють скорочення мишечних До. судин, інші — матки або серця. Спеціальні дії на тих, що діляться До. здійснюються при використанні радіації або цитостатічеських речовин, блокуючих ділення К. Іммунізация стимулює діяльність лімфоїдних До., що виробляють антитіла до чужорідних білок, запобігаючи тим самим багато захворювань.
Літ.: Кольцов Н. До., Організація клітки, М. — Л., 1936; Вільсон Е., Клітка і її роль в розвитку і спадковості, пер.(переведення) з англ.(англійський), т. 1—2, М. — Л., 1936—1940; Насонов Д. Н. і Александров Ст Я., Реакція живої речовини на зовнішні дії, М. — Л., 1940; Кедровський Би. Ст, Цитологія білкових синтезів в тваринній клітці, М., 1959; Мезія Д., Мітоз і фізіологія клітинного ділення, пер.(переведення) з англ.(англійський), М., 1963; Керівництво по цитології, т. 1—2, М. — Л., 1965—66; Бродський Ст Я., Трофіка клітки, М., 1966; Жива клітина, [Сб. ст.], пер.(переведення) з англ.(англійський), М., 1966; Де Робертіс Е., Новінський Ст, Саес Ф., Біологія клітки, пер.(переведення) з англ.(англійський), М., 1967; Васильев Ю. М. і Маленков А. Р., Клітинна поверхня і реакції кліток, Л., 1968; Алов І. А., Брауде А. І., Аспіз М. Е., Основи функціональної морфології клітки, 2 видавництва, М., 1969; Льові А., Сикевіц Ф., Структура і функції клітки, пер.(переведення) з англ.(англійський), М., 1971; Handbook of molecular cytology, ed. A. Lima-de-faria, Amst., 1969.