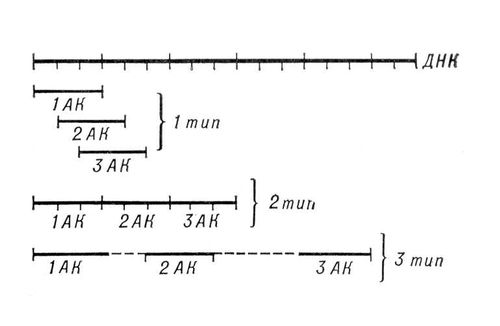
Генетичний код , система зашифровуваної спадкової інформації в молекулах нуклеїнових кислот, що реалізовується у тварин, рослин, бактерій і вірусів у вигляді послідовності нуклеотидів . У природних нуклеїнових кислотах — дезоксирибонуклеїновою (ДНК) і рібонуклеїновою (РНК) — зустрічаються 5 поширених типів нуклеотидів (по 4 в кожній нуклеїновій кислоті), що розрізняються по вхідній в їх склад азотистій підставі (див. Пуріновиє підстави, Пірімідіновиє підстави ). У ДНК(дезоксирибонуклеїнова кислота) зустрічаються підстави: аденін (А), гуанін (Г), цитозін (Ц), тимін (Т); у РНК(рибонуклеїнова кислота) замість тиміну присутній урацил (У). Окрім них, у складі нуклеїнових кислот виявлено близько 20 підстав, що рідко зустрічаються (т.з. неканонічних, або мінорних), а також незвичайних цукрів. Т. до. кількість кодуючих знаків Р. до. (4) і число різновидів амінокислот в білці (20) не збігаються, кодове число (тобто кількість нуклеотидів, що кодують 1 амінокислоту) не може дорівнювати 1. Різних поєднань по 2 нуклеотиди можливо лише 4 2 =16, але цього також недостатньо для зашифровуваної всіх амінокислот. Американський учений Г. Гамов запропонував (1954) модель триплетного Р. до., тобто такого, в якому 1 амінокислоту кодує група з трьох нуклеотидів наз.(назив) кодоном . Число можливих триплетів рівне 4 3 =64, а це більш ніж втричі перевищує число поширених амінокислот, у зв'язку з чим було висловлено припущення, що кожній амінокислоті відповідає декілька кодонів (т.з. вирожденность коди). Було запропоновано багато різних моделей Р. до., з яких на серйозну увагу заслуговували три моделі (см. мал.(малюнок) ): код без ком, код без ком, що не перекривається, і код з комами, що перекривається. У 1961 Ф. Крік (Великобританія) із співробітниками отримав підтвердження гіпотези триплетної коди, що не перекривається, без ком. Встановлені наступні основні закономірності, що стосуються Р. к.: 1) між послідовністю нуклеотидів і кодованою послідовністю амінокислот існує лінійна відповідність (колінеарність Р. до.); 2) прочитування Р. до. починається з певної крапки; 3) прочитування йде в одному напрямі в межах одного гена; 4) код є таким, що не перекривається; 5) при прочитуванні не буває проміжків (код без ком); 6) Р. до., як правило, є виродженим, тобто 1 амінокислоту кодують 2 і більш за триплети-синоніми (вирожденность Р. до. зменшує вірогідність того, що мутаційна заміна підстави в триплеті приведе до помилки); 7) кодове число рівне трьом; 8) код в живій природі універсальний (за деякими виключеннями). Універсальність Р. до. підтверджується експериментами по синтезу білка in vitro. Якщо в безклітинну систему, отриману з одного організму (наприклад, кишкової палички), додати нуклєїновокислотную матрицю, отриману з ін. організму, далеко віддаленого від першого в еволюційному відношенні (наприклад, проростков гороху), то в такій системі, йтиме білковий синтез. Завдяки роботам американських генетиків М. Ніренберга, С. Очоа, Х. Корана відомий не лише склад, але і порядок нуклеотидів у всіх кодонах (див. таблиці., побудовану за даними дослідів з кишковою паличкою).
З 64 кодонів у бактерій і фагов 3 кодони — УАА, УАГ і УГА — не кодують амінокислот; вони служать сигналом до звільнення поліпептидного ланцюга з рибосоми, тобто сигналізують про завершення синтезу поліпептиду. Їх назиают кодонами, що термінують. Існують також 3 сигнали про початок синтезу — це т.з. кодони, що ініціюють, — АУГ, ГУГ і УУГ, — які, будучи включеними на початку відповідної інформаційної РНК(рибонуклеїнова кислота) (І-РНК), визначають включення формілметіоніна в перше положення поліпептидного ланцюга, що синтезується. Приведені дані справедливі для бактерійних систем; для вищих організмів багато що ще не ясне. Так, кодон УГА у вищих організмів може бути значущим; не зовсім зрозумілий також механізм ініціації поліпептиду.
Реалізація Р. до. у клітці відбувається в два етапи. Перший з них протікає в ядрі; він носить назву транскрипції і полягає в синтезі молекул І-РНК на відповідних ділянках ДНК(дезоксирибонуклеїнова кислота). При цьому послідовність нуклеотидів ДНК(дезоксирибонуклеїнова кислота) «переписується» в нуклеотідную послідовність РНК(рибонуклеїнова кислота). Другий етап — трансляція — протікає в цитоплазмі, на рибосомах; при цьому послідовність нуклеотидів І-РНК переводиться в послідовність амінокислот в білці: цей етап протікає за участю транспортної РНК(рибонуклеїнова кислота) (Т-РНК) і відповідних ферментів (див. Білки, розділ Біосинтез).
Повний «словник» генетичної коди для амінокислот
Перша «буква»
Друга буква
Третя «буква»
В
Ц
А
Г
В
УУУ
Феніла- ланін
УЦУ
Серин
УАУ
Тирозин
УГУ
Цистин
В
УУЦ
УЦЦ
УАЦ
УГЦ
Ц
УУА
Лейцин
УЦА
УАА
Кінець синтезу
УГА
Кінець синтезу
А
УУГ*
УЦГ
УАГ
УГГ
Триптофан
Г
Ц
ЦУУ
Лейцин
ЦЦУ
Пролін
ЦАУ
Гістидин
ЦГУ
Аргінін
В
ЦУЦ
ЦЦЦ
ЦЯЦЬ
ЦГЦ
Ц
ЦУА
ЦЦА
ЦАА
Глутамін
ЦГА
А
ЦУГ
ЦЦГ
ЦАГ
ЦГГ
Г
А
АУУ
Ізолей- цин
АЦУ
Треонін
ААУ
Аспарагин
АГУ
Серин
В
АУЦ
АЦЦ
ААЦ
АГЦ
Ц
АУА
АЦА
ААА
Лізин
АГА
Аргінін
А
АУГ*
Метіонін
АЦГ
ААГ
АГГ
Г
Г
ГУУ
Валін
ГЦУ
Аланін
ГАУ
Аспараги- нова кислота
ГГУ
Гліцин
В
ГУЦ
ГЦЦ
ГАЦ
ГГЦ
Ц
ГУА
ГЦА
ГАА
Глутамі- нова кислота
ГГА
А
ГУГ*
ГЦГ
ГАГ
ГГГ
Г
* На початку ланцюга І-РНК даний кодон визначає початок синтезу поліпептидного ланцюга і кодує амінокислоту формілметіонін. Від готових поліплоїдних ланцюгів формільная група або вся амінокислота може бути відщеплена з допомогою відповідних ферментів.
Літ.: Загальна природа генетичної коди для білків, в збірці: Молекулярна генетика, пер.(переведення) з англ.(англійський), М., 1963; Крик Ф., Генетичний код (I), в кн.: Структура і функція клітки, пер.(переведення) з англ.(англійський), М., 1964, с. 9—23; Ніренберг М., Генетичний код (II), там же, с. 24—41; Хейс В., Генетика бактерій і бактеріофагів, пер.(переведення) з англ.(англійський), М., 1965; Хартман Ф., Саськайнд З., Дія гена, пер.(переведення) з англ.(англійський), М., 1966; Бреслер С. Е., Введення в молекулярну біологію, 2 видавництва, М. — Л., 1966; Інгрем Ст, Біосинтез макромолекул, пер.(переведення) з англ.(англійський), М., 1966; Лобашев М. Е., Генетика, 2 видавництва, Л., 1967; Уотсон Дж., Молекулярна біологія гена, пер.(переведення) з англ.(англійський), М., 1967; Сойфер Ст Н., Молекулярні механізми мутагенезу, М., 1969; Дубінін Н. П.. Загальна генетика, М., 1970.