Білки, протеїни, високомолекулярні природні органічні речовини, побудовані з амінокислот і що грають фундаментальну роль в структурі і життєдіяльності організмів. Саме Б. (ферменти і ін.) здійснюють обмін речовин і енергетичні перетворення, нерозривно пов'язані з активними біологічними функціями. Б. входять до складу складних клітинних структур — органел. І хоча органели містять також інші речовини (ліпіди,вуглеводи, нуклеїнові кислоти, неорганічні компоненти), Би. особливо важливі; вони — основні структуроутворювачі і грають провідну роль у виконанні фізіологічних функцій. Наприклад, завдяки відповідній організації різного роду Б. біологічні мембрани, покриваючі клітки, активно (з витратою енергії) переносять в клітку або з клітки певні молекули і іони. Зокрема, транспорт катіонів створює електричну поляризацію, необхідну для процесів збудження . У рухових апаратах — м'язових волокнах і інших — комплекси специфічних Би. здійснюють скорочення, перетворюючи хімічну енергію на механічну роботу. Діяльність Би. багато в чому пов'язана з різними небілковими речовинами, з яких найбільше біологічне значення мають нуклеїнові кислоти. Проте вирішальним чинником молекулярних механізмів всіх активних проявів життєдіяльності є Б. У цьому сенсі підтверджено і деталізовано відоме положення Ф. Енгельса о Б. як основі біологічної форми руху матерії (див. «Анті-Дюрінг», 1966, с. 78). Молекули Б. у структурному відношенні нескінченно всілякі — жорсткість і точність унікальної організації поєднуються в них з гнучкістю і пластичністю (див. нижче Структура Б.). Все це створює неозорі функціональні потенції; тому Б. і з'явилися тим винятковим матеріалом, який послужив основою виникнення життя на Землі. Б. — один з основних продуктів живлення людини і тварин (див. Білкові корми ), вони служать джерелом відновлення і оновлення цитоплазми кліток, утворення ферментів, гормонів і ін. (див. Білковий обмін ).
Физико-хімічні властивості Б. Молекули Б. мають масу від десятків тис. до 1 млн. і вище. Так, фермент рібонуклеаза має молекулярну масу 12 700, дихальний пігмент равлика гемоціанин — 6 600 000. Елементарний склад більшості Б.: 50,6—54,5% вуглецю, 6,5—7,3% водню, 21,5—23,5% кисню, 15—17,6% азоту, 0,3—2,5% сірки; до складу ряду Б. входить і фосфор. Відомості про молекулярну масу і ряд властивостей молекул Би. можна отримати, досліджуючи їх осадження (седиментацію) в ультрацентрифузі, дифузію, в'язкість, розчинність і светорассеяніє. Все Б. з дуже великою молекулярною масою побудовані з дрібніших часток — субодиниць. Розчинні Б. — гідрофільні колоїди, що активно зв'язують воду; їх розчини володіють значною в'язкістю, низьким осмотичним тиском. Молекули Б. не проходять через напівпроникні мембрани, володіють слабкою здібністю до дифузії. Б. — амфотерниє електроліти, т.к. імеют вільні карбоксильні (кислотні) і аміни (основні) групи. Ізоелектрична точка різних Би. неоднакова: для альбуміну плазми крові вона дорівнює 4,7, для зеїну кукурудзи 6,2. Би. мають електричний заряд, що змінюється залежно від структури Б. і реакції середовища. У електричному полі розчинені Би. рухаються (електрофорез ), причому напрям і швидкість руху неоднакові для різних Б. Растворімость Би. варіює не менше, ніж інші їх властивості. Одні Б. легко розчиняються у воді, інші вимагають для розчинення невеликих концентрацій солей, треті переходять в розчин лише під впливом сильних лугів і т.п. З розчинів Би. неоднаково осідають органічними речовинами (наприклад спиртами) або високими концентраціями солей (висолюються). Істотні відмінності в розчинності і інших властивостях використовуються при виділенні індивідуальних Би. з тих складних систем, в яких вони зустрічаються в природі. Після очищення багато Б. здатні кристалізуватися.
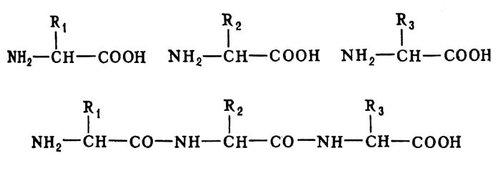
Структура Б. Білок всіх організмів складається з 20 видів амінокислот. Кожен Би. характеризується певним асортиментом і кількісним співвідношенням амінокислот. У молекулах Би. амінокислоти сполучені між собою пептидними зв'язками (—СО—NH—) в лінійній послідовності ( мал. 1 ), складовій так звану первинну структуру Б.
Амінокислотні (поліпептидні) ланцюги, що містять амінокислоту цистин, в місцях його розташування скріпляють дисульфідними зв'язками (—S—S—). Між амінокислотами в Би., як правило, не існує інших хімічних зв'язків, окрім пептидних і дисульфідних. Для кожного Б. не лише склад, але і послідовність амінокислот в поліпептидному ланцюзі — первинна структура — строго індивідуальні; будь-яка ланка ланцюга — сповна певна амінокислота. Всі багаточисельні види Б., що існують в природі, розрізняються по первинній структурі; потенційне можливе їх число практично необмежено. Індивідуальна первинна структура кожного Б. зберігається в поколіннях завдяки точній передачі відповідної спадкової інформації (див. нижче Біосинтез Би.). Для аналізу первинної структури Б. розроблені спеціальні методи. При переварюванні певними ферментами, наприклад трипсином, кожен Би. дає свій набір фрагментів (пептидів). При відповідному їх розділенні на аркуші паперу виходить «пептидна карта», яка, подібно до відбитку пальця, характерна для даного Б. Разделеніє на пептиди і визначення будови кожного з них в окремості — основна дорога розшифровки первинної структури Б.
Окрім пептидних і дисульфідних зв'язків, в молекулі Б. є багаточисельні зв'язки з меншою енергією взаємодії, що мають велике значення для внутрішньої організації і функції Б. Среді цих зв'язків найбільш істотні так звані гідрофобні зв'язки, що створюються неполярними бічними групами амінокислот. Ці групи, позбавлена спорідненість, до води мають тенденцію контактувати між собою усередині молекули Б. Кроме того, в молекулі Б. є водневі зв'язки, що утворюються полярними групами, наприклад —СО—NH—, а також електростатичні взаємодії між групами, що несуть електричні заряди.
Просторова конфігурація (конформація) поліпептидного ланцюга Б. визначається його первинною структурою і умовами середовища. За звичайних умов (температура не вище 40°С, нормальний тиск і т.д.) Би. характеризуються внутрішньомолекулярною впорядкованістю. «Хребет» поліпептидного ланцюга
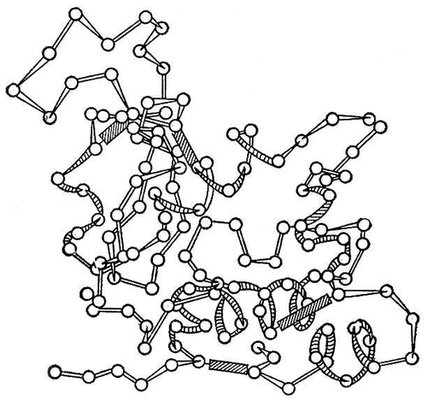
місцями може закручуватися спіраллю або утворювати повністю витягнуті відрізки (вторинна структура). У обох випадках виникає система водневих зв'язків. Але в значній частині «хребта» геометрична регулярність може бути відсутньою. Поліпептидний ланцюг в цілому «упаковується» і жорстко фіксується за допомогою взаємодій бічних груп амінокислот (третинна структура). Залежно від укладання поліпептидних ланцюгів форма молекул Би. варіює від фіблярної (витягнутою, ниткоподібною) до глобулярної (округлою). Детальна конфігурація глобулярних молекул складна і своєрідна для кожного Б. У молекулі превалює досконала впорядкованість, що поширюється на положення окремих атомів. Проте деякі периферичні ділянки можуть бути закріплені менш жорстко, а занурені в розчинник гідрофільні бічні групи залишаються сповна гнучкими. Конформація деяких Би., наприклад лізоциму ( мал. 2 ), розкрита рентгеноструктурнимі дослідженнями. Створення впорядкованої міцної конформації Б. визначається цілими системами взаємодій, що знаходяться у взаємній залежності. Зміни конформації Б., середовища, що викликаються змінами, або реакціями, в які Б. вступають, пов'язані із зміною ряду взаємодій. Конформаційні переходи охоплюють молекулу Б. цілком або обмежуються певними районами. При нагріванні, різкому підкисленні середовища і інших сильних діях відбувається «плавлення» молекули Б. — перехід в стан безладного клубка. Це, як правило, спричиняє за собою ряд інших перетворень, загальний результат яких позначають як денатурацію Б. (див. Біополімери ) . При цьому знижується розчинність Би., посилюється в'язкість їх розчинів втрачаються ферментативні і інші біологічні властивості.
Кожен з незліченної безлічі тих, що існують Би. має особливу спадково детерміновану первинну структуру, властиву лише йому. Це обумовлює строго індивідуальну систему внутрішньомолекулярних зв'язків, тобто унікальну конформацію Б. Поетому кожен Би. характеризується власною «хімічною топографією» і своєрідними поєднаннями хімічних груп, що просторово зближують. Частина таких поєднань служить функціональними центрами молекул Б. Благодаря структурній відповідності, що нагадує відношення ключа до замку (комплементу), функціональні центри «взнають» і вибірково приєднують речовини, на яких відповідних Би. «встановлені». Функціональні — активні центри Би.-ферментов специфічно приєднують субстрати і активують їх, прискорюючи і направляючи хімічні перетворення. За допомогою особливих центрів взаємного скріплення («контактних майданчиків») визначені Б. з'єднуються по декількох разом (структура 4-го порядку) або створюють значно складніші системи (самосборка крупних білкових структур). Процеси самосборки істотні для морфогенезу .
Вивчення структури Б. дає можливість переходити до їх синтезу. У 1955 була з'ясована структура інсуліну, молекула якого складається з двох порівняно коротких поліпептидних ланцюгів (21 і 30 амінокислотних залишків). Услід за цим була розкрита первинна структура гемоглобіну, рібонуклеази, трипсину і ряду інших Би. ( мал. 3 ). Шляхом хімічного синтезу спочатку були отримані складні пептиди з властивостями гормонів, потім удалося синтезувати гормон інсулін, нарешті — фермент рібонуклеазу. Правильність хімічної формули інсуліну і рібонуклеази підтвердилася тим, що синтетичні Б. не відрізнялися від Би., продукованих організмом, ні по физико-хімічних властивостях, ні по біологічній активності. Встановлена повністю або частково первинна структура понад 200 Би.
Класифікація Б. До цих пір немає єдиного принципу класифікації Б. Прі діленні всіх відомих Би. на групи враховують і їх склад (будова), физико-хімічні властивості (розчинність, лужність), походження і роль в організмі. Б. ділять на простих — протеїни, що складаються лише з амінокислот, і складні — протеїди, до складу молекули яких входять, окрім амінокислот, і інші з'єднання. До простих Би. відносяться альбумін, глобуліни, гистони, глутеліни, проламіни,протаміни і протєїноїди . До складних Би. відносяться глікопротеїди (що містять, окрім амінокислот, вуглеводи), ліпопротєїди (ліпіди, що містять), нуклеопротєїди (у їх склад входять і нуклеїнові кислоти), фосфопротєїди (фосфорні, що містять кислоти) і хромопротєїди (пігментні металовмісні групи, що мають).
В. А. Беліцер .
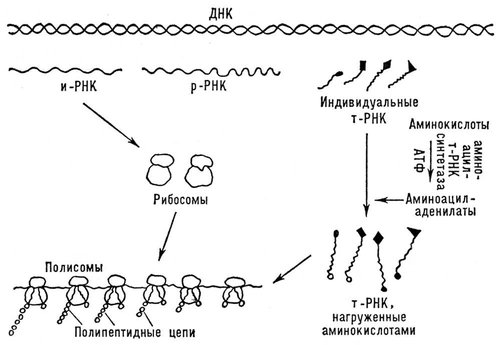
Біосинтез Би. — процес освіти Б. з амінокислот в клітках живих організмів. З'ясування механізму цього процесу, що має величезне біологічне значення, можна віднести до найважливіших досягнень науки 20 ст Біосинтез Би. йде за допомогою особливих складних механізмів, що забезпечують впорядковане відтворення специфічних Би. унікальної структури. Механізми ці єдині або вельми схожі для найрізноманітніших кліток і організмів, в них беруть участь нуклеїнові кислоти, особливо рибонуклеїнові кислоти (РНК). Цей процес йде з використанням енергії, накопиченій у вигляді аденозинтрифосфорної кислоти (АТФ) (див. Біоенергетика ).
Біосинтез Би. відбувається на особливих рібонуклеопротєїдних частках — рибосомах, що складаються з майже рівних кількостей рибосомної РНК(рибонуклеїнова кислота) ( р -РНК) і білків. Первинна структура (послідовність амінокислот) поліпептидних ланцюжків, що синтезуються, забезпечується з'єднанням з рибосомами особою матричною, або інформаційної, рібонуклеїнової кислоти (І-РНК, або М-КОД-РНК), яка містить інформацію про специфічну будову Б., «закодовану» у вигляді послідовного розташування нуклеотидів, складових І-РНК. Цю інформацію І-РНК отримує від дезоксирибонуклеїнової кислоти (ДНК), що зберігає і передавальною її по спадку. Амінокислоти, перш ніж попасти в рибосоми, активуються, отримуючи енергію від АТФ і утворюючи з'єднання з аденіловою кислотою. (Активовані амінокислоти є змішаним ангідридом амінокислоти і аденілової кислоти — аміноациладенілат.) Далі, залишок даної амінокислоти переноситься на відповідну транспортну рибонуклеїнову кислоту (Т-РНК). Обидва ці процесу каталізують одним і тим же ферментом (аміноациладенілатсинтетазой або АМІНОАЦИЛТ-РНК-СИНТЕТАЗОЙ), специфічним для кожної амінокислоти. Певній амінокислоті відповідають одна або декілька специфічних для неї Т-РНК. Все Т-РНК порівняно нізкополімерни, містять близько 80 нуклеотідних залишків. Вони побудовані по загальному плану: на початку ланцюга знаходиться 5-гуаніловая кислота, а в кінці — угрупування, що часто обмінюється, з двох залишків цитидилової кислоти і аденозіна, до якого і приєднується залишок амінокислоти. Залишок амінокислоти, сполучений з Т-РНК, далі переноситься на рибосоми, де і відбувається утворення поліпептидного ланцюжка Б. ( мал. 4 ). Т. о., рибосомна стадія — центральний етап біосинтезу Б. В процесі біосинтезу Б. рибосоми з'єднуються в ланцюжки за допомогою І-РНК, утворюючи активні белоксинтезірующие структури —полірібосоми, або полісоми.
І-РНК синтезується на матриці ДНК(дезоксирибонуклеїнова кислота). У унікальній послідовності нуклеотидів ДНК(дезоксирибонуклеїнова кислота) лінійно «записана» генетична інформація про послідовність амінокислотних залишків в поліпептидних ланцюжках Би.
В новоутвореній І-РНК виходить нуклеотідная послідовність, відповідна матричною ДНК(дезоксирибонуклеїнова кислота), — послідовність комплементу, яка визначає первинну структуру поліпептидного ланцюжка, що синтезується. Включення кожній амінокислоти обумовлюється (кодується) певними групами з трьох нуклеотідних залишків (триплетами). Кожній амінокислоті відповідає декілька триплетів, або кодонів, для яких тепер встановлені склад і послідовність нуклеотидів (див. Генетичний код ).
В полісомах Т-РНК, навантажена амінокислотою, приєднується до відповідних кодонів І-РНК. Це приєднання здійснюється усередині рибосоми через взаємодію підстав комплементу: аденіна з урацилом або тиміном і гуаніна з цитозіном. При цьому Т-РНК приєднується до кодону триплетом комплементу, що міститься в ній, званим антикодоном. У міру просування рибосоми по нуклеотідной ланцюжку І-РНК до сусідніх кодонів приєднуються нові молекули Т-РНК, навантажені амінокислотами. Попередня Т-РНК при цьому звільняється, приєднуючи свою амінокислоту карбоксильним кінцем до аміногрупи нової амінокислоти з утворенням пептидного зв'язку. Т. о., поліпептидний ланцюжок зростає у міру просування рибосоми по І-РНК і звільняється після закінчення свого синтезу, пройдя відповідну ділянку І-РНК, комплемент даному структурному гену (цистрону ) ДНК(дезоксирибонуклеїнова кислота).
Процес біосинтезу Б. не вичерпується освітою поліпептидних ланцюжків, тобто створенням первинної структури Б. Далєє відбувається згортання ланцюжків в спіралі, їх «укладання» і взаємодія, і освіта вторинною, третинною і, інколи, четвертинної структури. Проте можливо, що приведена схема не вичерпує всіх доріг біосинтезу Б.
Вельми важлива проблема регуляції біосинтезу Б., що визначає включення або виключення синтезу тих або інших Би. під впливом внутрішніх (у тому числі диференціювання кліток і тканин) або зовнішніх імпульсів і що створює умови для синтезу Б. у даній диференційованій клітці.
Теоретична і експериментальна розробка проблеми біосинтезу Б. має не лише найважливіше теоретичне, але і практичне значення, оскільки, відкриваючи підходи до дії на цей процес, вона намічає дороги лікування ряду захворювань, а також впливу на продуктивність багатьох сільськогосподарських рослин і тварин.
У зв'язку з важливим значенням Би. розробляються нові методи здобуття Б. і амінокислот шляхом промислового мікробіологічного синтезу, тобто вирощуванням мікробів (наприклад, дріжджів і ін.) на дешевій сировині (наприклад, нафті, газі і ін.).
І. Б. Збарський .
Літ.: Волькенштейн М. Ст, Молекули і життя, М., 1965, гл.(глав) 3—5; Гауровіц Ф., Хімія і функції білків, пер.(переведення) з англ.(англійський), [2 видавництва], М., 1965; Біосинтез білка і нуклеїнових кислот, під ред. А. С. Спіріна, М., 1965; Сисакян Н. М. і Гладілін До. Л., Біохімічні аспекти синтезу білка, в кн.: Успіхи біологічної хімії, т. 7, М., 1965, с. 3; Молекули і клітки. [Сб. ст.], пер.(переведення) з англ.(англійський), М., 1966, с. 7—27, 94—106; Шамін А. Н., Розвиток хімії білка, М., 1966; Введення в молекулярну біологію, пер.(переведення) з англ.(англійський), М., 1967.