Біологічні мембрани, тонкі пограничні структури молекулярних розмірів, розташовані на поверхні кліток і субклітинних часток, а також канальців і бульбашок, пронизливих протоплазму. Товщина Б. м. не перевищує 100 . Найважливіша функція Б. м. — регулювання транспорту іонів, цукрів, амінокислот і інших продуктів обміну речовин (див. Проникність біологічних мембран ) . Спочатку термін «Би. м.» використовували при описі всіх видів пограничних структур, що зустрічаються в живому організмі, — покривних тканин, слизистих оболонок шлунку і кишечника, стінок кровоносних судин і ниркових канальців, мієлінових оболонок нервових волокон, оболонок еритроцитів і ін. До середини 20 ст було доведено, що в більшості пограничних структур ефективну бар'єрну функцію виконують не всі елементи цих складних утворень, а лише мембрани кліток. За допомогою електронного мікроскопа і рентгеноструктурного аналізу удалося показати спільність будови поверхневих клітинних мембран еритроцитів, нервових і м'язових клітин, бактерій, плазмалеми рослинних кліток і ін. з мембранами субклітинних структур — ендоплазматичній мережі, мітохондрій, клітинних ядер, лізосом, хлоропластів і ін. Б.м. займають величезну площу (наприклад, в організмі людини лише поверхневі мембрани мають площу, рівну десяткам тис. м 2 ) і грають універсальну регуляторну роль в обміні речовин. Тому вивчення структури і функцій Би. м. — одне з найважливіших завдань цитології і молекулярної біології. Функції Б. м. багатообразні (див. таблиці.).
Деякі функції біологічних мембран
Функція
Вигляд мембрани
Активний транспорт речовин
Загальна і виборча дифузія невеликих молекул і іонів
Регулювання транспорту іонів і продуктів метаболізму усередині кліток
Всі види мембран
Електроізоляційні властивості
Мієлін
Генерація нервового імпульсу
Мембрани нервових клітин
Перетворення світлової енергії в хімічну енергію аденозинтрифосфорної кислоти (АТФ)
Мембрани хлоропластів
Перетворення енергії біологічного окислення в хімічну енергію макроергічних фосфатних зв'язків в молекулі аденозинтрифосфорної кислоти (АТФ)
Мембрани мітохондрій
Фагоцитоз, піноцитоз, антигенні властивості
Мембрани спеціалізованих кліток
Покриваючи клітку і відокремлюючи її від довкілля, Би. м. забезпечують морфологічну цілісність кліток і субклітинних часток, їх міцність і еластичність. Підтримуючи нерівномірний розподіл іонів калія, натрію, хлору і ін. між протоплазмою і що оточує середовищем, вони сприяють появі різниці біоелектричних потенціалів . Властивості Б. м. в значній мірі визначають генерування і проведення збудження як в нервових і м'язових клітинах, так і в місцях контакту між ними, тобто в синаптічеських закінченнях (див. Синапси ) . Би. м. мітохондрій служать местомом строго впорядкованого розташування ферментів, що беруть участь в синтезі багатих енергією з'єднань.
Функціональні властивості Б. м. тісно пов'язані з їх структурною організацією і в значній мірі визначаються нею. Ще в 1902, вивчаючи проникність клітинних мембран, німецький учений Е. Овертон відмітив, що через мембрани найлегше проникають речовини, добре розчинні в ліпідах, і передбачив наявність останніх в поверхневій клітинній мембрані. У 1926 американські біологи Е. Гортер і Ф. Грендел виділили з гемолізованих еритроцитів людини ліпіди і розташували їх у вигляді мономолекулярного шару на поверхні води; загальна площа цього шару приблизно в 2 рази перевищувала поверхню еритроцитів. З цього вони зробили вивід, що ліпіди Б. м. розташовані у вигляді бімолекулярного шару. Поверхневе натягнення клітинної мембрани (0,1 мн/м-код, або дінів/см ) менше натягнення шару чистого ліпіду (10 мн/м-код, або дінів/см ) і близько до поверхневого натягнення білків. Тому було передбачено, що в Би. м. бімолекулярний ліпідний шар покритий з двох сторін шарами білка (структура «сендвіча»). Вивчення клітинної поверхні за допомогою поляризаційного мікроскопа дозволило передбачити, що молекули ліпідів розташовані перпендикулярно, а молекули білка — паралельно клітинній поверхні. Методом електропровідності удалося виміряти електричну ємкість клітинної мембрани, рівну 1 мкф/см 2 , і розрахувати товщину її ліпідного шару, яка виявилася рівною 55 . На основі всіх цих даних англійські біологи Л. Данієллі і Г. Даусон в 1935 запропонували модель Би. м., в основних межах що задовольняє сучасним уявленням про структуру Б. м.
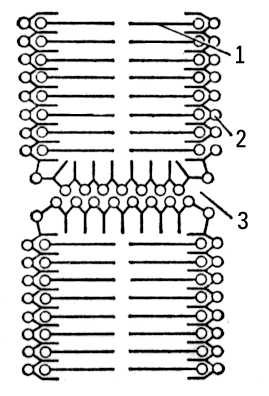
Методами рентгеноструктурного аналізу, електронній мікроскопії, а також оптичними і біохімічними методами показано, що поверхнева клітинна мембрана і мембрани субклітинних часток — мітохондрій, ядер, мікросом, лізосом і ін. — мають схожу структуру. Вони складаються з бімолекулярного ліпідного шару (в основному з фосфоліпідів ) завтовшки 35 і двох неліпідних шарів завтовшки 20 кожен (американський дослідник Дж. Робертсон). Зовнішня поверхня багато Б. м. покрита мукополісахарідамі . Внутрішня поверхня Б. м. вистилає структурним або ферментним білком ( мал. 1 , 2 ). Передбачається, що між молекулами фосфоліпідів і білка існує електростатичне тяжіння. Мембрани мітохондрій декілька відрізняються по структурі від поверхневої клітинної мембрани ( мал. 3 ). Мабуть, фосфоліпіди і білки у складі внутрішньої мембрани мітохондрій зв'язані між собою міцною гідрофобною взаємодією і утворюють комплекси («одиниці, що повторюються»), з яких побудована вся мембрана.
Значний прогрес в уявленнях про структуру і функцію Б. м. досягнуть при вивченні їх моделей — штучних фосфоліпідних мембран, що складаються з бімолекулярного шару фосфоліпідів. Фізичні властивості такої плівки близькі до властивостей природних Би. м.: товщина її досягає 61, а електрична ємкість 1 мкф/см 2 . При додаванні в розчин, що омиває штучну мембрану, невеликої кількості білка електричний опір її різко зменшується (~ у 1000 разів), наближаючись до електричного опору природних Би. м. За певних умов в такій «реконструйованій» мембрані можуть виникати електричні коливання, по амплітуді, тривалості і умовам виникнення електричні коливання, що нагадують, в нервовому волокні при збудженні. Додавання в розчин, що омиває цю мембрану, антибіотиків типа валіноміцина, Граміцидину і ін. викликало появу виборчої проникності для іонів калія і натрію. Дослідження Б. м. ведуться інтенсивно; в найближчому майбутньому можна чекати повної розшифровки їх структури і функції.
Літ.: Керівництво по цитології, т. 1, М-код.—Л., 1965, гл.(глав) 2; Робертіс Е. де, Новінський Ст, Саес Ф., Біологія клітки, пер.(переведення) з англ.(англійський), М., 1967; Робертсон Дж., Мембрана живої клітини, в збірці: Структура і функція клітки, пер.(переведення) з англ.(англійський), М., 1964; Finean J. Ст, The molecular organization of cell membranes, «Progress in Biophysics and Molecular Biology», 1966, v. 16, р. 143—70.