Проникність біологічних мембран, найважливіша властивість біологічних мембран (БМ), що полягає в їх здатності пропускати в клітку і з неї різні метаболіти (амінокислоти, цукру, іони і т.п.). П. би. м. має велике значення для осморегуляції і підтримки постійності складу клітки, її физико-хімічний гомеостаз ; грає важливу роль в генерації і проведенні нервового імпульсу, в енергозабезпеченні клітки, сенсорних механізмах і ін. процесах життєдіяльності. П. би. м. обумовлена особливостями будови БМ, що є осмотичним бар'єром між кліткою і середовищем, і служить характерним прикладом єдності і взаємозв'язку між структурою і функцією на молекулярному рівні.
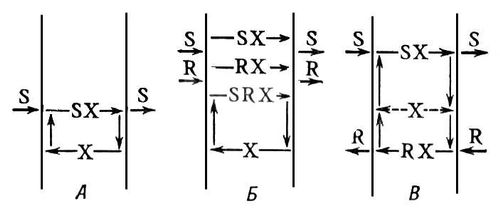
БМ проникні лише для невеликого числа низькомолекулярних жиророзчинних речовин (гліцерин, спирти, сечовина і ін.). Така проникність (проста дифузія) грає порівняно малу роль в процесах перенесення речовин через мембрани. Важливіші процеси перенесення (транслокації) речовин через БМ відбуваються за участю специфічних систем транспорту. Передбачають, що ці системи містять мембранні переносники (білки або ліпопротєїди) і, можливо, ряд ін. компонентів, що здійснюють пов'язані з транспортом функції (наприклад, рецепторні). Переносник (або їх система) зв'язує переносиму речовину (субстрат) і може переміщатися в мембрані. Якщо переносники непорушно фіксовані в БМ, то вважають, що в БМ існують специфічні для переносимої речовини пори або канали ( мал. 1 ). Якщо переносник зв'язується з субстратом шляхом невалентних взаємодій (іонними, гідрофобними і ін. силами), то такий процес називається вторинною транслокацією; розрізняють 3 її типи ( мал. 2 ): полегшена дифузія (уніпорт), котранспорт (симпорт) і протівотранспорт (антипорт). Механізм полегшеної дифузії не залежить від перенесення ін. речовин в клітку або з клітки. Цим способом переноситься, наприклад, глюкоза в еритроцити. Котранспорт — спільний транспорт двох (або більш) речовин в одному напрямі. Так, транспорт глюкози і амінокислот через слизисті оболонки тонкого кишечника зв'язаний з транспортом іонів Na + . Механізм протівотранспорта має на увазі сполучення перенесення речовини в одному напрямі з потоком ін. речовини в протилежному напрямі. Цим способом здійснюється протилежно направлене перенесення іонів Na + і К + в нервових клітинах (див. Мембранна теорія збудження ). Процеси зв'язаного транспорту (симпорт і антипорт) мають велике значення в тих випадках, коли переносима речовина рухається проти градієнта концентрації (з області меншої в область більшої концентрації). Такий активний транспорт, на відміну від пасивного транспорту (по концентраційному градієнту), вимагає витрат енергії. Енергозабезпечення активного транспорту досягається за рахунок сполучення вторинної транслокації з ферментативними реакціями розриву або утворення хімічних зв'язків. При цьому енергія хімічного перетворення витрачається на підтримку осмотичного потенціалу або асиметрії по обидві сторони мембрани.
Транспорт речовин через БМ, пов'язаний з розривом або утворенням валентних зв'язків, називається первинною транслокацією. Типовий приклад такого процесу — робота «натрієвого насоса», зв'язана з хімічною реакцією гідролізу багатого енергією аденозинтрифосфату (АТФ), що каталізує ферментом аденозінтріфосфатазой. Гідроліз АТФ супроводиться перенесенням іонів Na + з клітки і вступом в клітку іонів К + ; передбачають, що переносником іонів К + є вільний фермент, а іонів Na + — фосфорильований фермент, що утворюється в ході гідролізу АТФ. До цих пір не удалося виділити переносників з БМ кліток тварин. У бактерій чітко доведено (головним чином генетичними методами) існування переносників — т.з. пермеаз, деякі з них (наприклад, м-код-білок — переносник лактози в кишкової палички) виділені в чистому вигляді. Є дані, що показують, що активний транспорт цукрів і амінокислот у бактерій зв'язаний з окисленням d-молочної комітети. У деяких бактерій виявлено велике число «зв'язуючих білків», які, можливо, є рецепторними компонентамі відповідних транспортних систем.
П. би. м. регулюється гормонами і ін., біологічно активними речовинами. Так, деякі стероїдні гормони, інсулін і ін. збільшують проникність мембран еритроцитів, м'язових і жирових кліток. П. би. м. збудливих кліток (наприклад, нервових) залежить від особливих речовин — медіаторів (ацетілхолін і ін.). На П. би. м. для іонів сильно впливають антибіотики (валіноміцин, Граміцидин, нонактін), а також деякі синтетичні поліефіри. У дослідженнях П. би. м. — одній з найважливіших проблем молекулярній біології — велике значення мають модельні мембрани: ліпідні моношари, штучні двошарові мембрани, багатошарові замкнуті мембрани (ліпосоми) і т.п. Для вивчення П. би. м. широко застосовуються електро-хімічні, фізичні і хімічні методи. Див. також Біологічні мембрани .
Літ.: Біологічні мембрани, М., 1973; Гершановіч Ст Н., Біохімічні і генетичні основи перенесення вуглеводів в бактерійну клітку, М., 1973; Никольський Н. Н. Трошин А. С., Транспорт Цукрів через клітинні мембрани, Л., 1973; Ташмухамедов Би. А., Гагельганс А. І., Активний транспорт іонів через біологічні мембрани, Таш., 1973; Mitchell P., Translocations through natural membranes, «Advances in Enzymology and Related Areas of Molecular Biology», 1967, v. 29; Kaback Н. R., Transport, «Annual Review of Biochemistry», 1970, v. 39.