Біополімери , високомолекулярні природні з'єднання, що є структурною, основою всіх живих організмів і що грають визначальну роль в процесах життєдіяльності. ДО Б. відносяться білки, нуклеїнові кислоти і полісахариди; відомі також змішані Б. — глікопротеїди, ліпопротєїди, гліколіпіди і ін.
Біологічні функції Б. Нуклеїнові кислоти виконують в клітці генетичні функції. Послідовність мономірних ланок (нуклеотидів) в дезоксирибонуклеїновій кислоті — ДНК(дезоксирибонуклеїнова кислота) (інколи в рибонуклеїновій кислоті — РНК(рибонуклеїнова кислота)) визначає (у формі генетичної коди ) послідовність мономірних ланок (амінокислотних залишків) у всіх білках, що синтезуються, і, т. о., будова організму і що протікають в йому біохімічні процеси. При діленні кожної клітки обидві дочірні клітки отримують повний набір генів завдяки попередньому самоудвоєнію (реплікації ) молекул ДНК(дезоксирибонуклеїнова кислота). Генетична інформація з ДНК(дезоксирибонуклеїнова кислота) переноситься на РНК(рибонуклеїнова кислота), що синтезується на ДНК(дезоксирибонуклеїнова кислота) як на матриці (транскрипція ) . Ета т.з. інформаційна РНК(рибонуклеїнова кислота) (І-РНК) служить матрицею при синтезі білка, що відбувається на особливих органоїдах клітки — рибосомах (трансляція ) за участю транспортної РНК(рибонуклеїнова кислота) (Т-РНК). Біологічна мінливість, необхідна для еволюції, здійснюється на молекулярному рівні за рахунок змін в ДНК(дезоксирибонуклеїнова кислота) (див. Мутація ).
Білки виконують в клітці ряд найважливіших функцій. Білки-ферменти здійснюють всі хімічні реакції обміну речовин в клітині, проводячи їх в необхідній послідовності і з потрібною швидкістю. Білки м'язів, джгутиків мікробів, клітинних ворсинок і ін. виконують скоротливу функцію, перетворюючи хімічну енергію на механічну роботу і забезпечуючи рухливість організму в цілому або його частин. Білки — основний матеріал більшості клітинних структур (в т.ч. в спеціальних видах тканин) всіх живих організмів, оболонок вірусів і фагов. Оболонки кліток є ліпопротєїднимі мембранами, рибосоми побудовані з білка і РНК(рибонуклеїнова кислота) і т.д. Структурна функція білків тісно пов'язана з регуляцією вступу різних речовин в субклітинні органели (активний транспорт іонів і ін.) і з ферментативним каталізом. Білки виконують і регуляторні функції (репрессори ), «забороняючи» або «вирішуючи» прояв того або іншого гена. У вищих організмах є білки — переносники тих або інших речовин (наприклад, гемоглобін — переносник молекулярного кисню) і імунні білки, що захищають організм від чужорідних речовин, проникаючих в організм (див. Імунітет ) . Полісахариди виконують структурну, резервну і деякі інші функції. Білки і нуклеїнові кислоти утворюються в живих організмах шляхом матричного ферментативного біосинтезу . Є тепер і біохімічні системи позаклітинного синтезу Б. за допомогою ферментів, виділених з кліток. Розроблені методи хімічного синтезу білків і нуклеїнових кислот.
Первинна структура Б. Состав і послідовність мономірних ланок Би. визначають їх т.з. первинну структуру. Всі нуклеїнові кислоти є лінійними гетерополімерами — сахарофосфатнимі ланцюжками, до ланкам яких приєднані бічні групи — азотисті підстави: аденін і тимін (у РНК(рибонуклеїнова кислота) — урацил), гуанін і цитозін; в деяких випадках (головним чином в Т-РНК) бічні групи можуть бути представлені іншими азотистими підставами. Білки — також гетерополімери; молекули їх утворені однією або декількома поліпептидними ланцюжками, сполученими дисульфідними містками. До складу поліпептидних ланцюгів входить 20 видів різних мономірних ланок — залишків амінокислот. Молекулярна маса ДНК(дезоксирибонуклеїнова кислота) варіює від декількох млн. (в дрібних вірусів і бактеріофагів) до ста млн. і більш (в крупніших фагов); бактерійні клітки містять по одній молекулі ДНК(дезоксирибонуклеїнова кислота) з молекулярною масою в декілька млрд. ДНК(дезоксирибонуклеїнова кислота) вищих організмів може мати і велику молекулярну масу, але виміряти її доки не удалося із-за розривів в молекулах ДНК(дезоксирибонуклеїнова кислота), що виникає при їх виділенні. Рибосомні РНК(рибонуклеїнова кислота) мають молекулярну масу від 600 тис. до 1,1 млн., інформаційна (І-РНК) — від сотень тисяч до декількох мільйонів, транспортна (Т-РНК) — близько 25 тис. Молекулярна маса білків варіює від 10 тис. (і менш) до мільйонів; у останньому випадку, проте, зазвичай можливе розділення білкової частки на субодиниці, сполучені між собою слабкими, переважно гідрофобними, зв'язками.
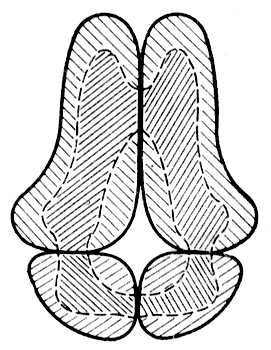
Конформація, тобто та або інша просторова форма молекул Би., визначається їх первинною структурою. Залежно від хімічної будови і зовнішніх умов молекули Б. можуть знаходитися або в одній або в декількох переважних конформаціях (нативні стани Б.:, що зазвичай зустрічаються в природних умовах, наприклад, глобулярна будова білків, подвійна спіраль ДНК(дезоксирибонуклеїнова кислота)), або приймати багато більш менш рівноімовірних конформацій. Білки ділять по просторовій структурі на фіблярних (ниткоподібні) і глобулярних; білки-ферменти, білки-переносники, імунні і деякі інші мають, як правило, глобулярну структуру. Для ряду білків — гемоглобін, міоглобін, лізоцим, рібонуклеаза і ін. — ця структура встановлена у всіх деталях (з визначенням за допомогою рентгеноструктурного аналізу розташування кожного атома). Вона визначається послідовністю амінокислотних залишків і утворюється і підтримується відносно слабкими взаємодіями між мономірними ланками поліпептидних ланцюгів у водно-сольовому розчині (кулонівські і дипольні сили, водневі зв'язки, гідрофобні взаємодії), а також дисульфідними зв'язками. Глобула білка формується так, що більшість полярних гідрофільних амінокислотних залишків виявляються зовні і контактують з розчинником, а більшість неполярних (гідрофобних) залишків знаходяться усередині і ізольовано від взаємодії з водою. Молекули білка, що володіють надлишком неполярних груп, коли частина з них виявляється на поверхні глобули, утворюють вищу, т.з. четвертинну структуру, при якій декілька глобул агрегують, взаємодіючи між собою в основному неполярними ділянками ( мал. 1 ). Просторова структура кожного білка-ферменту унікальна і забезпечує необхідне для його функціонування розташування в просторі всіх ланок Би., особливо т.з. активних центрів . В той же час вона не абсолютно жорстка і допускає необхідні в процесі функціонування (при взаємодії з субстратами, інгібіторами і іншими речовинами) конформаційні зрушення і зміни.
Просторова структура нативної ДНК(дезоксирибонуклеїнова кислота) утворена двома нитками комплементу і є подвійну спіраль Крику — Уотсона; у ній протилежні азотисті підстави попарно зв'язані водневими зв'язками — аденін з тиміном і гуанін з цитозіном. Стійкість подвійної спіралі забезпечується, поряд з водневими зв'язками, також гідрофобною взаємодією між плоскими кільцями азотистих підстав, розташованих стопкою (взаємодія стопки, або стакинг). Нитки РНК(рибонуклеїнова кислота) спіралізують лише частково. ДНК(дезоксирибонуклеїнова кислота) вірусів бактеріофагів, бактерій, а також мітохондріальна у ряді випадків є замкнутим кільцем; при цьому поряд із спіраллю Крику — Уотсона спостерігається ще додаткова т.з. надспіралізує.
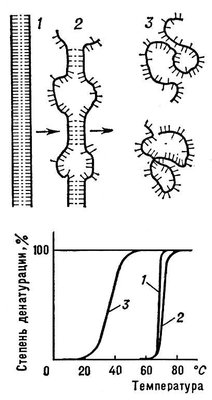
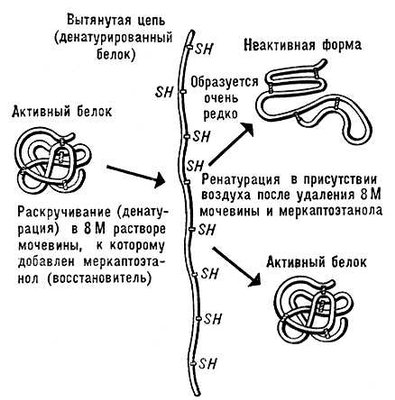
Денатурація Б. Нарушеніє нативної просторової структури Б. при різних діях (підвищення температури, зміна концентрації металів, кислотності розчину і ін.) називається денатурацією і у ряді випадків оборотно (зворотний процес називається ренатурацією; мал. 2 ). Молекули Б. — кооперативні системи: поведінка їх залежить від взаємодій складників. Кооперативність молекул Би. визначається тим, що повороти окремих ланок із-за внутрішньомолекулярних взаємодії залежать від конформації сусідніх ланок. У основі денатурації Б. при зміні зовнішніх умов зазвичай лежать кооперативні конформаційні перетворення (наприклад, переходи а-спіраль — b-структура, а-спіраль — клубок, b-структура — клубок для поліпептидів, перехід глобула — клубок для глобулярних білків, перехід спіраль — клубок для нуклеїнових кислот). На відміну від фазових переходів (кипіння рідини, плавлення кристала), що є граничним випадком кооперативних процесів і що відбуваються стрибком, кооперативні переходи Б. здійснюються в кінцевому, хоча і порівняно вузькому, інтервалі змін зовнішніх умов. У цьому інтервалі одновимірні, лінійні молекули (нуклеїнові кислоти поліпептиди), що зазнають перехід спіраль — клубок, розбиваються на спіральних, що чергуються, і клубкообразниє ділянки ( мал. 3 ).
Перехід спіраль — клубок в ДНК(дезоксирибонуклеїнова кислота) спостерігається при підвищенні температури, додаванні в розчин кислоти або лугу, а також під впливом інших денатуруючих агентів. Цей перехід в гомополінуклеотідах відбувається при нагріванні в інтервалі десятих доль °С, у фагових і бактерійної ДНК(дезоксирибонуклеїнова кислота) — в інтервалі 3—5°С ( мал. 3 ), в ДНК(дезоксирибонуклеїнова кислота) вищих організмів — в інтервалі 10—15 °С. Чим вище гетерогенність ДНК(дезоксирибонуклеїнова кислота), тим ширше інтервал переходу і менше здатність молекул ДНК(дезоксирибонуклеїнова кислота) до ренатурації. Перехід спіраль — клубок в різних видах РНК(рибонуклеїнова кислота) носить менш кооперативний характер ( мал. 4 ) і відбувається в ширшому інтервалі температурних або інших денатуруючих дій.
Би. — полімерні електроліти, їх просторова конформація і кооперативні переходи залежать як від міри іонізації молекули, так і від концентрації іонів в середовищі, що впливає на електростатичні взаємодії як між окремими частинами молекули, так і між Би. і розчинником.
Будова і біологічні функції Б. Будова Б. — результат тривалої еволюції на молекулярному рівні, унаслідок чого ці молекули ідеально пристосовані до виконання своїх біологічних завдань. Між первинною структурою, конформацією Б. і конформаційними переходами, з одного боку, і їх біологічними функціями — з іншою, існують тісні зв'язки, дослідження яких — одне з головних завдань молекулярній біології . Встановлення таких зв'язків в ДНК(дезоксирибонуклеїнова кислота) дозволило зрозуміти основні механізми реплікації,транскрипції і трансляції, а також мутагенезу і деяких інших найважливіших біологічних процесів. Лінійна структура молекули ДНК(дезоксирибонуклеїнова кислота) забезпечує запис генетичної інформації, її подвоєння при матричному синтезі ДНК(дезоксирибонуклеїнова кислота) і здобуття (також шляхом матричного синтезу) багатьох копій з одного і того ж гена, тобто молекул І-РНК. Сильні ковалентні зв'язки між нуклеотидами забезпечують збереження генетичної інформації при всіх цих процесах. В той же час відносний слабкі зв'язки між нитками ДНК(дезоксирибонуклеїнова кислота) і можливість обертання довкола простих хімічних зв'язків забезпечують гнучкість і лабільність просторової структури, необхідні для розділення ниток при реплікації і транскрипції, а також рухливість молекули І-РНК, матрицею, що служить, при біосинтезі білка (трансляція). Дослідження просторової структури і конформаційних змін білків-ферментів на різних стадіях ферментативної реакції при взаємодії з субстратами і коферментами дає можливість встановити механізми біокаталізу і зрозуміти природу величезного прискорення хімічних реакцій, здійснюваного ферментами.
Методи дослідження Б. Прі дослідженні будови і конформаційних перетворень Би. широко використовуються як очищені природні Б., так і їх синтетичні моделі, які простіше по будові і легше піддаються дослідженню. Так, при вивченні білків моделями служать гомогенні або гетерогенні поліпептиди (із заданим або випадковим чергуванням амінокислотних залишків). Моделями ДНК(дезоксирибонуклеїнова кислота) і РНК(рибонуклеїнова кислота) є відповідні синтетичні гомогенні або гетерогенні полінуклеотиди. До методів дослідження Б. і їх моделей відносяться рентгеноструктурний аналіз, електронна мікроскопія, вивчення спектрів поглинання, оптичній активності, люмінесценції, методи светорассеянія і динамічного подвійного променезаломлення, седиментаційний метод, віскозиметрія физико-хімічні методи розділення і очищення і ряд ін. Всі методи, розроблені для вивчення синтетичних полімерів, застосовні і до Б. Прі трактуванню властивостей Би. і їх моделей, закономірностей їх конформаційних перетворень використовуються також методи теоретичної фізики (статистичної фізики, термодинаміки, квантової механіки і ін.).
Літ.: Бреслер С. Е., Введення в молекулярну біологію, М-код.—Л., 1966; Волькенштейн М. Ст, Молекули і життя, М., 1965; Уотсон Дж., Молекулярна біологія гена, пер.(переведення) з англ.(англійський), М., 1967; фізичні методи дослідження білків і нуклеїнових кислот, М., 1967.