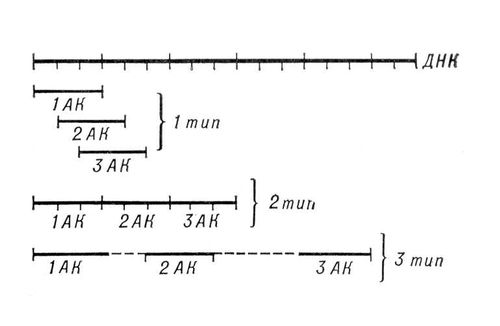
Генетический код, система зашифровки наследственной информации в молекулах нуклеиновых кислот, реализующаяся у животных, растений, бактерий и вирусов в виде последовательности нуклеотидов. В природных нуклеиновых кислотах— дезоксирибонуклеиновой (ДНК) и рибонуклеиновой (РНК) — встречаются 5 распространённых типов нуклеотидов (по 4 в каждой нуклеиновой кислоте), различающихся по входящему в их состав азотистому основанию (см. Пуриновые основания, Пиримидиновые основания). В ДНК(дезоксирибонуклеиновая кислота) встречаются основания: аденин (А), гуанин (Г), цитозин (Ц), тимин (Т); в РНК(рибонуклеиновая кислота) вместо тимина присутствует урацил (У). Кроме них, в составе нуклеиновых кислот обнаружено около 20 редко встречающихся (т. н. неканонических, или минорных) оснований, а также необычных сахаров. Т. к. количество кодирующих знаков Г. к. (4) и число разновидностей аминокислот в белке (20) не совпадают, кодовое число (т. е. количество нуклеотидов, кодирующих 1 аминокислоту) не может быть равно 1. Различных сочетаний по 2 нуклеотида возможно лишь 42=16, но этого также недостаточно для зашифровки всех аминокислот. Американский учёный Г. Гамов предложил (1954) модель триплетного Г. к., т. е. такого, в котором 1 аминокислоту кодирует группа из трёх нуклеотидов, наз.(назыв) кодоном. Число возможных триплетовравно 43=64, а это более чем втрое превышает число распространённых аминокислот, в связи с чем было высказано предположение, что каждой аминокислоте соответствует несколько кодонов (т. н. вырожденность кода). Было предложено много различных моделей Г. к., из которых серьёзного внимания заслуживали три модели (см. рис.(рисунок)): перекрывающийся код без запятых, неперекрывающийся код без запятых и код с запятыми. В 1961 Ф. Крик (Великобритания) с сотрудниками получил подтверждение гипотезы триплетного неперекрывающегося кода без запятых. Установлены следующие основные закономерности, касающиеся Г. к.: 1) между последовательностью нуклеотидов и кодируемой последовательностью аминокислот существует линейное соответствие (колинеарность Г. к.); 2) считывание Г. к. начинается с определённой точки; 3) считывание идёт в одном направлении в пределах одного гена; 4) код является неперекрывающимся; 5) при считывании не бывает промежутков (код без запятых); 6) Г. к., как правило, является вырожденным, т. е. 1 аминокислоту кодируют 2 и более триплетов-синонимов (вырожденность Г. к. уменьшает вероятность того, что мутационная замена основания в триплете приведёт к ошибке); 7) кодовое число равно трём; 8) код в живой природе универсален (за некоторыми исключениями). Универсальность Г. к. подтверждается экспериментами по синтезу белка in vitro. Если в бесклеточную систему, полученную из одного организма (например, кишечной палочки), добавить нуклеиновокислотную матрицу, полученную из др. организма, далеко отстоящего от первого в эволюционном отношении (например, проростков гороха), то в такой системе, будет идти белковый синтез. Благодаря работам американских генетиков М. Ниренберга, С. Очоа, Х. Корана известен не только состав, но и порядок нуклеотидов во всех кодонах (см. табл., построенную по данным опытов с кишечной палочкой).
Из 64 кодонов у бактерий и фагов 3 кодона — УАА, УАГ и УГА — не кодируют аминокислот; они служат сигналом к освобождению полипептидной цепи с рибосомы, т. е. сигнализируют о завершении синтеза полипептида. Их назыают терминирующими кодонами. Существуют также 3 сигнала о начале синтеза — это т. н. инициирующие кодоны — АУГ, ГУГ и УУГ, — которые, будучи включенными в начале соответствующей информационной РНК(рибонуклеиновая кислота) (и-РНК), определяют включение формилметионина в первое положение синтезируемой полипептидной цепи. Приведённые данные справедливы для бактериальных систем; для высших организмов многое ещё не ясно. Так, кодон УГА у высших организмов может быть значащим; не совсем понятен также механизм инициации полипептида.
Реализация Г. к. в клетке происходит в два этапа. Первый из них протекает в ядре; он носит название транскрипции и заключается в синтезе молекул и-РНК на соответствующих участках ДНК(дезоксирибонуклеиновая кислота). При этом последовательность нуклеотидов ДНК(дезоксирибонуклеиновая кислота) «переписывается» в нуклеотидную последовательность РНК(рибонуклеиновая кислота). Второй этап — трансляция— протекает в цитоплазме, на рибосомах; при этом последовательность нуклеотидов и-РНК переводится в последовательность аминокислот в белке: этот этап протекает при участии транспортной РНК(рибонуклеиновая кислота) (т-РНК) и соответствующих ферментов (см. Белки, раздел Биосинтез).
Полный «словарь» генетического кода для аминокислот
Первая «буква»
Вторая буква
Третья «буква»
У
Ц
А
Г
У
УУУ
Фенила- ланин
УЦУ
Серин
УАУ
Тирозин
УГУ
Цистин
У
УУЦ
УЦЦ
УАЦ
УГЦ
Ц
УУА
Лейцин
УЦА
УАА
Конец синтеза
УГА
Конец синтеза
А
УУГ*
УЦГ
УАГ
УГГ
Триптофан
Г
Ц
ЦУУ
Лейцин
ЦЦУ
Пролин
ЦАУ
Гистидин
ЦГУ
Аргинин
У
ЦУЦ
ЦЦЦ
ЦАЦ
ЦГЦ
Ц
ЦУА
ЦЦА
ЦАА
Глутамин
ЦГА
А
ЦУГ
ЦЦГ
ЦАГ
ЦГГ
Г
А
АУУ
Изолей- цин
АЦУ
Треонин
ААУ
Аспарагин
АГУ
Серин
У
АУЦ
АЦЦ
ААЦ
АГЦ
Ц
АУА
АЦА
ААА
Лизин
АГА
Аргинин
А
АУГ*
Метионин
АЦГ
ААГ
АГГ
Г
Г
ГУУ
Валин
ГЦУ
Аланин
ГАУ
Аспараги- новая кислота
ГГУ
Глицин
У
ГУЦ
ГЦЦ
ГАЦ
ГГЦ
Ц
ГУА
ГЦА
ГАА
Глутами- новая кислота
ГГА
А
ГУГ*
ГЦГ
ГАГ
ГГГ
Г
* В начале цепи и-РНК данный кодон определяет начало синтеза полипептидной цепи и кодирует аминокислоту формилметионин. От готовых полиплоидных цепей формильная группа или вся аминокислота может быть отщеплена с помощью соответствующих ферментов.
Лит.: Общая природа генетического кода для белков, в сборнике: Молекулярная генетика, пер.(перевод) с англ.(английский), М., 1963; Крик Ф., Генетический код (I), в кн.: Структура и функция клетки, пер.(перевод) с англ.(английский), М., 1964, с. 9—23; Ниренберг М., Генетический код (II), там же, с. 24—41; Хэйс У., Генетика бактерий и бактериофагов, пер.(перевод) с англ.(английский), М., 1965; Хартман Ф., Саскайнд З., Действие гена, пер.(перевод) с англ.(английский), М., 1966; Бреслер С. Е., Введение в молекулярную биологию, 2 изд., М. — Л., 1966; Ингрэм В., Биосинтез макромолекул, пер.(перевод) с англ.(английский), М., 1966; Лобашев М. Е., Генетика, 2 изд., Л., 1967; Уотсон Дж., Молекулярная биология гена, пер.(перевод) с англ.(английский), М., 1967; Сойфер В. Н., Молекулярные механизмы мутагенеза, М., 1969; Дубинин Н. П.. Общая генетика, М., 1970.