Ядро клітинне, обов'язкова, поряд з цитоплазмою, складова частина клітки у простих, багатоклітинних тварин і рослин, що містить хромосоми і продукти їх діяльності. По наявності або відсутності в клітках Я. всі організми ділять на еукаріот і прокаріот . В останніх немає оформленого Я. (відсутня його оболонка), хоча дезоксирибонуклеїнова кислота (ДНК) є. У Я. зберігається основна частина спадкової інформації клітки; гени, що містяться в хромосомах, грають головну роль в передачі спадкових ознак у ряді кліток і організмів. Я. знаходиться в постійній і тісній взаємодії з цитоплазмою; у нім синтезуються молекули-посередники, що переносять генетичну інформацію до центрів білкового синтезу в цитоплазмі. Т. о. Я. управляє синтезами всіх білків і через них — всіма фізіологічними процесами в клітці. Тому отримувані експериментально без'ядерні клітки і фрагменти кліток завжди гинуть; при пересадці Я. у такі клітки їх життєздатність відновлюється. Я. вперше спостерігав чеський учений Я. Пуркине (1825) в яйцеклітині курки; у рослинних клітках Я. описав англійський учений Р. Броун (1831—33), в тваринних клітках — німецький учений Т. Шванн (1838—39).
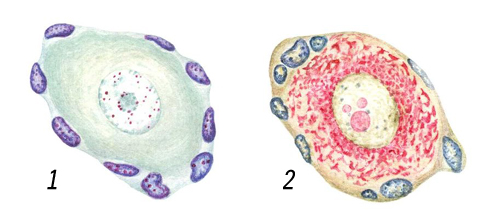
Зазвичай Я. у клітці одне, знаходиться поблизу її центру, має вигляд сферичної або еліпсоїдної бульбашки ( фігури 1—3, 5, 6 ). Рідше Я. буває неправильної ( фігура 4 ) або складної форми (наприклад, Я. лейкоцитів, макронуклеуси інфузорій). Нерідкі двух- і багатоядерні клітки, що зазвичай утворюються шляхом ділення Я. без ділення цитоплазми або шляхом злиття декілька одноядерних кліток (т.з. симпласти, наприклад поперечнополосатиє м'язові волокна). Розміри Я. варіюють від ~ 1 мкм (в деяких простих) до ~ 1 мм (деякі яйцеклітини).
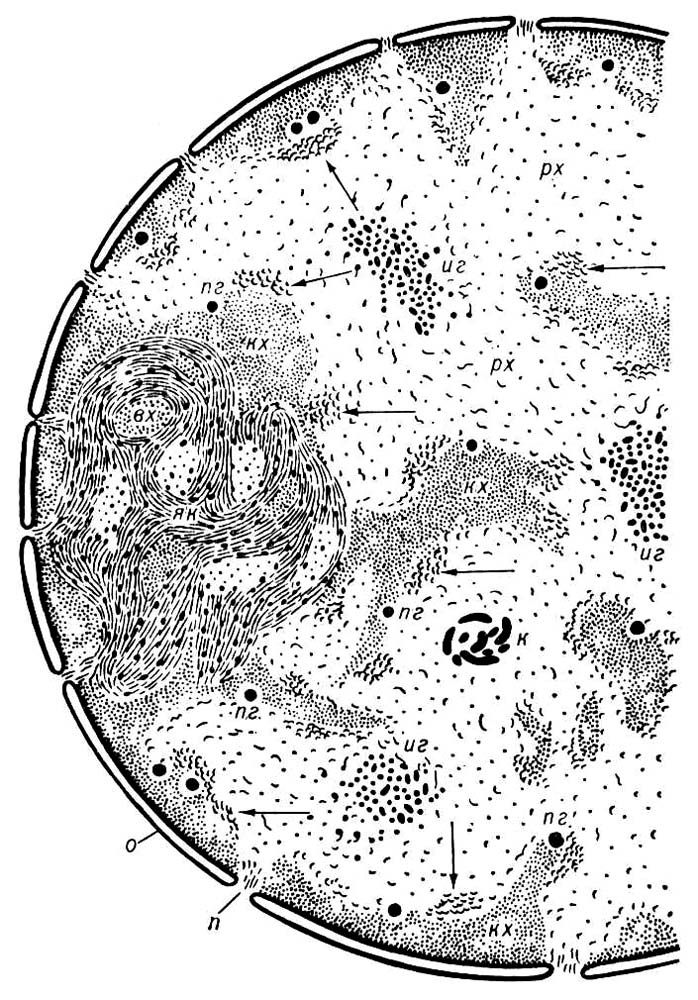
Я. відокремлено від цитоплазми ядерною оболонкою (ЯО), що складається з 2 паралельних ліпопротєїдних мембран завтовшки 7—8 нм , між якими знаходиться вузький перинуклеарний простір. ЯО пронизана порами діаметром 60—100 нм , на краях яких зовнішня мембрана ЯО переходить у внутрішню. Частота пір різна в різних клітках: від одиниць до 100—200 на 1 мкм 2 поверхні Я. По краю пори розташовується кільце щільного матеріалу — так званий аннулус. У просвіті пори часто є центральна гранула діаметром 15—20 нм , сполучена з аннулусом радіальною волокниною. Разом з порою ці структури складають поровий комплекс, який, мабуть, регулює проходження макромолекул через ЯО (наприклад, вхід в Я. білкових молекул, вихід з Я. рібонуклеопротєїдних часток і т. п.). Зовнішня мембрана ЯО місцями переходить в мембрани ендоплазматичній мережі ; вона зазвичай несе белоксинтезірующие частки — рибосоми . Внутрішня мембрана ЯО інколи утворює впячиванія в глиб Я. Содержімоє Я. представлено ядерним соком (каріолімфой, каріоплазмою) і зануреними в нього оформленими елементами — хроматином, ядерцями і ін. Хроматин — це більш менш розпушений в тому, що не ділиться Я. матеріал хромосом, комплекс ДНК(дезоксирибонуклеїнова кислота) з білками — так званий дезоксирібо-нуклеопротеїд (ДНП). Він виявляється за допомогою кольорової реакції Фельгена на ДНК(дезоксирибонуклеїнова кислота) ( фігури 1 і 8 ). При діленні Я. (див. Мітоз ) весь хроматин конденсується в хромосоми; після закінчення мітоза велика частина ділянок хромосом знову розпушується; ці ділянки (так званий еухроматин) містять в основному унікальні (що не повторюються) гени. Інші ділянки хромосом залишаються щільними (так званий гетерохроматін); у них розташовуються послідовності ДНК(дезоксирибонуклеїнова кислота), що головним чином повторюються. У тому, що не ділиться Я. велика частина еухроматину представлена рихлою мережею фібрил ДНП завтовшки 10 — 30 нм , гетерохроматін — щільними глибкамі (хромоцентрамі), в яких та ж волокнина щільно упакована. Частина еухроматину також може переходити в компактний стан; такий еухроматин вважається неактивним відносно синтезу РНК(рибонуклеїнова кислота). Хромоцентри зазвичай граничать з ЯО або ядерцем. Є дані про те, що волокнина ДНП закріплена на внутрішній мембрані ЯО.
В тому, що не ділиться Я. відбувається синтез (реплікація ) ДНК(дезоксирибонуклеїнова кислота), що вивчається шляхом реєстрації включених в Я. мічених радіоактивними ізотопами попередників ДНК(дезоксирибонуклеїнова кислота) (зазвичай тимідину). Показано, що по довжині хроматинової волокнини є безліч ділянок (так званих репліконов), кожен зі своєю точкою початку синтезу ДНК(дезоксирибонуклеїнова кислота), від якої реплікація поширюється в обидві сторони. Унаслідок реплікації ДНК(дезоксирибонуклеїнова кислота) подвоюється і самі хромосоми.
В хроматині Я. відбувається прочитування закодованої в ДНК(дезоксирибонуклеїнова кислота) генетичної інформації шляхом синтезу на ДНК(дезоксирибонуклеїнова кислота) молекул матричною, або інформаційною, РНК(рибонуклеїнова кислота) (див. Транскрипція ), а також молекул інших типів РНК(рибонуклеїнова кислота), що беруть участь в білковому синтезі. Спеціальні ділянки хромосом (і відповідно хроматину) містять гени, що повторюються, які кодують молекули рибосомної РНК(рибонуклеїнова кислота); у цих місцях Я. формуються багаті рібонуклеопротєїдамі (РНП) ядерця , основна функція яких — синтез РНК(рибонуклеїнова кислота), що входить до складу рибосом. Поряд з компонентамі ядерця в Я. є і інші види часток РНК(рибонуклеїнова кислота). До них відноситься періхроматіновиє волокнина завтовшки 3—5 нм і періхроматіновиє гранули (ПГ) діаметром 40—50 нм , розташовані на кордонах зон рихлого і компактного хроматину. І ті та інші, ймовірно, містять матричну РНК(рибонуклеїнова кислота) в з'єднанні з білками, а ПГ відповідають її неактивній формі; спостерігався вихід ПГ з Я. у цитоплазму через пори ЯО. Є також інтерхроматіновиє гранули (20—25 нм ), а інколи і товсті (40—60 нм ) нитки РНП, скручені в клубки. У ядрах амеб є нитки РНП, скручені в спіралі (30—35 нм х 300 нм ); спіралі можуть виходити в цитоплазму і, ймовірно, містять матричну РНК(рибонуклеїнова кислота). Поряд з ДНК(дезоксирибонуклеїнова кислота) - і РНК(рибонуклеїнова кислота) -содержащимі структурами деякі Я. містять чисто білкові включення у вигляді сфер (наприклад, в Я. зростаючих яйцеклітин багатьох тварин, в Я. ряду простих), пучків волокнини або кристалоїдів (наприклад, в ядрах багато тканинних кліток тварин і рослин, макронуклеусах ряду інфузорій). У Я. виявлені також фосфоліпіди, ліпопротепди, ферменти (ДНК-полімераза, РНК(рибонуклеїнова кислота) -полімераза, комплекс ферментів оболонки Я., у тому числі аденозінтріфосфатаза, і ін.).
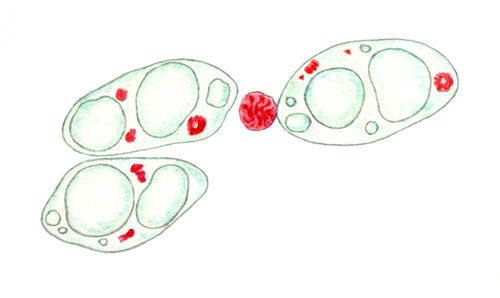
В природі зустрічаються різні спеціальні типи Я.: гігантські Я. зростаючих. яйцеклітин, особливо риб і земноводних; Я., гігантські політенні хромосоми, що містять (див. Політенія ), наприклад в клітках слинних залоз двокрилих комах; компактні, позбавлені ядерц Я. сперматозоїдів і мікронуклеуси інфузорій, суспіль заповнені хроматином і що не синтезують РНК(рибонуклеїнова кислота); Я., у яких хромосоми постійно конденсують, хоча ядерця утворюються (в деяких простих, у ряді кліток комах); Я., у яких сталося дву- або багатократне збільшення числа наборів хромосом (поліплоїдія ; фігури 7, 9 ).
Основний спосіб ділення Я. — мітоз, що характеризується подвоєнням і конденсацією хромосом, руйнуванням ЯО (виключення — багато простих і гриби) і правильною розбіжністю сестринських хромосом в дочірні клітки. Проте Я. деяких спеціалізованих кліток, особливо поліплоїдниє, можуть ділитися простою перешнуровуванням (див. Амітоз ) . Високополіплоїдниє Я. можуть ділитися не лише на 2, але і на багато частин, а також розмножуватися брунькуванням ( фігура 7 ). При цьому може відбуватися розділення цілих хромосомних наборів (т.з. сегрегація геномів).
Літ.: Керівництво по цитології, т. 1, М. —Л., 1965; Райків І. Б., Каріология простих Л., 1967; Робертіс Е., Новінський Ст, Саес Ф.,. Біологія клітки, пер.(переведення) з англ.(англійський), М., 1973; Ченцов Ю. С., Поляків Ст Ю., Ультраструктура клітинного ядра, М., 1974; The nucleus, ed. A. J. Dalton, F, Haguenau, N. Y. — L., 1968; The cell nucleus, ed. Н. Busch, v. 1—3, N. Y. — L., 1974.