Хромосоми (від кульгавий... і сома ) , органоїди клітинного ядра, сукупність яких визначає основні спадкові властивості кліток і організмів. Повний набір Х. у клітці, характерний для даного організму, називається каріотипом . В будь-якій клітці тіла більшості тварин і рослин кожна Х. представлена двічі: одна з них отримана від батька, інша — від матері при злитті ядер статевих кліток в процесі запліднення. Такі Х. називаються гомологічними, набір гомологічних Х. — диплоїдним. У хромосомному наборі кліток раздельнополих організмів присутня пара (або декілька пар) статевих хромосом, як правило, що розрізняються в різної полови по морфологічних ознаках; останні Х. називаються аутосомами. У ссавців в статевих Х. локалізовані гени, що визначають пів організму; у плодової мушки дрозофіли пів визначається співвідношенням статевих хромосом і аутосом (балансова теорія визначення підлоги).
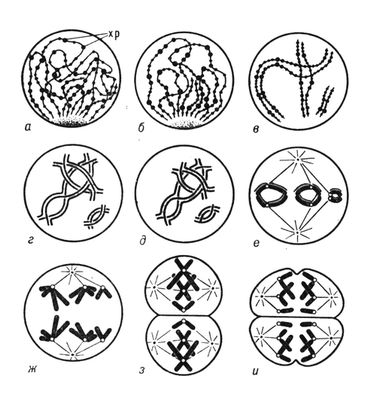
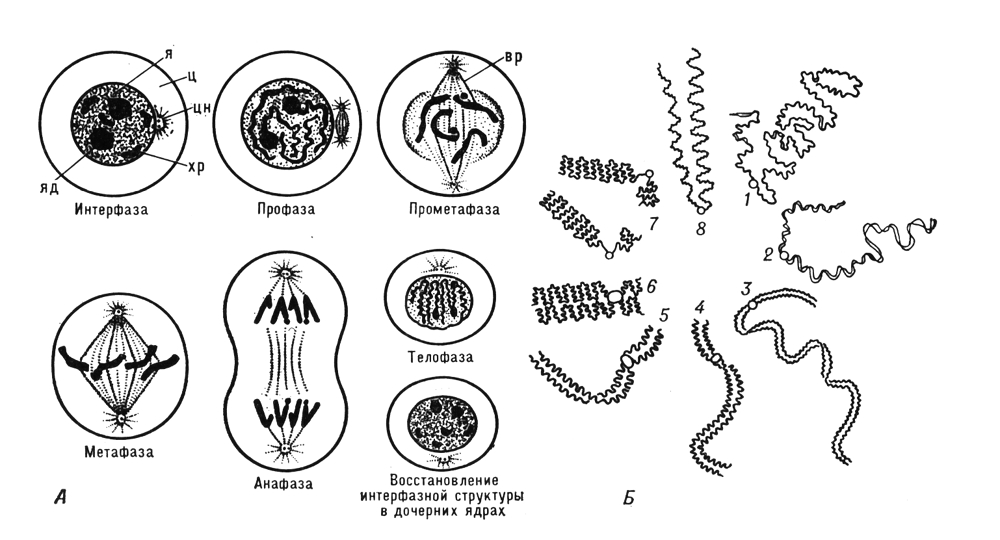
Спочатку Х. були описані як що інтенсивно забарвлюються основними фарбниками щільні тільця (німецьке вчене Ст Вальдейер, 1888). Проте виявилось, що зовнішній вигляд Х. істотно міняється на різних стадіях клітинного циклу, і як компактні утворення з характерною морфологією Х. чітко помітні в світловому мікроскопі лише в період клітинного ділення — в метафазі мітоза і мейозу ( мал. 1 , 2 ). Основу Х. на всіх стадіях клітинного циклу складають хромонеми — ниткоподібні структури, які під час ділення клітки щільно закручені, обумовлюючи спіралізує хромосом, а в клітці, що не ділиться, розкручені (деспіралізовани). При завершенні ділення клітки що розійшлися до її полюсів Х. розпушуються і оточуються ядерною мембраною. У період між двома діленнями клітки (ця стадія клітинного циклу називається інтерфазою ) деспіралізация Х. продовжується і вони стають малодоступними для спостереження в світловий мікроскоп. Морфологія Х. еукаріот істотно відрізняється від такої в прокаріот і вірусів. Прокаріоти (доядерні) і віруси містять зазвичай одну лінійну або кільце Х., яка не має надмолекулярної укладання і не відокремлена від цитоплазми ядерною оболонкою. Поняття Х. до генетичного апарату прокаріот застосовно лише умовно, т.к. оно сформувалося при вивченні Х. еукаріот і має на увазі наявність в Х. не лише складного комплексу біополімерів (нуклеїнових кислот і білків), але і специфічної надмолекулярної структури. Тому нижче дається опис лише Х. еукаріот. Зміни зовнішнього вигляду Х. у клітинному і життєвому циклах обумовлені особливостями функціонування Х. Общий же принцип їх організації, індивідуальність і безперервність Х. у ряді клітинних поколінь і організмів зберігаються незмінними. Докази тому отримані при біохімічному, цитологичеськом і генетичному дослідженнях Х. різних організмів. Вони лягли в основу хромосомній теорії спадковості .
Молекулярні основи будови Х. Значеніє Х. як клітинних органоїдів, відповідальних за зберігання, відтворення і реалізацію спадкової інформації, визначається властивостями біополімерів, що входять в їх склад. Перша молекулярна модель Х. була запропонована в 1928 Н. До. Кольцовим, що передбачив принципи їх організації. Запис спадкової інформації в Х. забезпечується будовою молекули дезоксирибонуклеїнової кислоти (ДНК), її генетичним кодом . В Х. зосереджено близько 99% всієї ДНК(дезоксирибонуклеїнова кислота) клітки, остання частина ДНК(дезоксирибонуклеїнова кислота) знаходиться в інших клітинних органоїдах, визначаючи цитоплазматичну спадковість. ДНК(дезоксирибонуклеїнова кислота) в Х. еукаріот знаходиться в комплексі з основними білками — гистонамі і з негистоновимі білками, які забезпечують складну упаковку ДНК(дезоксирибонуклеїнова кислота) в Х. і регуляцію її здібності до синтезу рибонуклеїнових кислот (РНК) — транскрипції .
Х. у інтерфазі. Х. виконує свої основні функції — репродукцію і транскрипцію — в інтерфазі, тому будова Х. на цій стадії клітинного циклу представляє особливий інтерес. У інтерфазі Х. погано помітні тому, що у зв'язку з активним синтезом РНК(рибонуклеїнова кислота) багато ділянок Х. (т.з. еухроматин ) сильно розкручені; інші ж (гетерохроматін ) не беруть участь в синтезі РНК(рибонуклеїнова кислота) і продовжують зберігати щільну упаковку (див. також Хромоцентр ) . В еухроматінових ділянках, окрім елементарних дезоксирібонуклеопротєїдних ниток (ДНП), є рібонуклеопротєїдниє частки діаметром 200—500, звані РНП-грануламі, інтергрануламі і періхроматіновимі гранулами. Ці частки є формою упаковки РНК(рибонуклеїнова кислота), синтезованою на Х. і сполученою з білком, і служать для завершення освіти інформаційною РНК(рибонуклеїнова кислота) і перенесення її в цитоплазму.
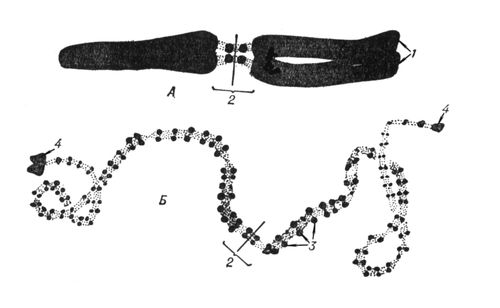
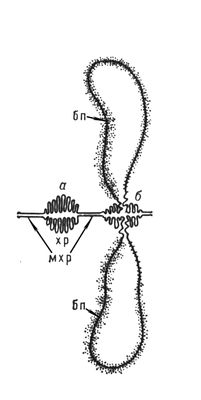
Для вивчення інтерфазних Х. використовують або біохімічні методи виділення речовини Х. — хроматину і розділення його на еухроматин і гетерохроматін або електронно-мікроскопічне дослідження інтактних ядер і ізольованого хроматину; як моделі інтерфазних Х. використовують гігантські Х. типа лампових щіток з ооцитов тваринних і багатонитчастих (політенні) Х. двокрилих. У Х. типа лампових щіток неактивні ділянки мають вигляд щільно упакованих структур — хромомер ( мал. 2 , 3 ), які виявляються і в Х. соматичних кліток, особливо в профазі мітоза, і розглядаються як характерні морфологічні, а можливо і функціональні, одиниці Х. У ділянках Х., що активно синтезують РНК(рибонуклеїнова кислота), хромомери розкручуються і утворюють бічні петлі, в яких молекули РНК(рибонуклеїнова кислота), з'єднуючись з білком, утворюють рібонуклеопротєїди (РНП) — частки, що є формою упаковки генних продуктів і що розрізняються в окремих бічних петлях по розмірах і морфологічним ознакам. У політенних Х., що виникають в тканинах двокрилих і деяких рослин за рахунок багатократної реплікації (подвоєння) початкової Х. без подальшої розбіжності дочерніх Х., неактивні ділянки мають форму дисків, а активні утворюють здуття — пуфи . В пуфах, так само як і в Х. типа лампових щіток, містяться частки РНП діаметром 200—500 . Електронно-мікроскопічні і біохімічні дослідження показали, що і в хроматині, виділеному з кліток, і в інтактних ядрах, і в гігантських Х. основною структурною одиницею є дезоксирібонуклеопротєїдная нитка (ДНП) діаметром,100—200 .
Вивчення політенних Х. у різних тканинах і на різних стадіях розвитку двокрилих показало, що число і набір активних пуфів мають тканинну і видову специфічність. Це означає, що хоча всі клітки багатоклітинного організму мають однаковий набір генів, лінійно розташованих в кожній Х., набір активних і неактивних в синтезі РНК(рибонуклеїнова кислота) ділянок Х. розрізняється в кожному типові кліток і на різних стадіях розвитку, тобто одна і та ж ділянка знаходиться в одних тканинах в еухроматічеськом, в інших — в гетерохроматичному стані. Окремі ділянки Х. знаходяться в гетерохроматичному поляганні в інтерфазі різних типів кліток; як правило, вони відрізняються присутністю послідовностей, що високоповторюються ДНК(дезоксирибонуклеїнова кислота). Постійно функціонує в інтерфазі всіх типів кліток є ядерцевий організатор — ділянка Х., де зосереджені гени рибосомної РНК(рибонуклеїнова кислота). У цій області формується ядерце, яке довго вважали самостійним органоїдом клітки. Воно є местомом формування попередників рибосом .
Х. у інтерфазном ядрі відокремлені від цитоплазми ядерною мембраною; багатьма ділянками (перш за все, теломерамі і центромерами ) вони сполучені з нею, завдяки чому, як вважають, кожна Х. займає в ядрі певне місце. При підготовці клітки до ділення в інтерфазі відбувається подвоєння Х. Каждая Х. будує свою копію на основі напівконсервативної реплікації ДНК(дезоксирибонуклеїнова кислота). Особливістю Х. еукаріот є існування багатьох крапок почала і завершення реплікації (в прокаріот лише одна крапка почала і одна точка завершення реплікації). Цим забезпечується можливість неодночасної реплікації різних ділянок Х. в ході синтетичного періоду і регуляція активності Х.
Х. в період мітоза і мейозу. Під час переходу клітки до ділення синтез ДНК(дезоксирибонуклеїнова кислота) і РНК(рибонуклеїнова кислота) в Х. припиняється, Х. набувають усе більш щільної упаковки (наприклад, в одній Х. людини ланцюжок ДНК(дезоксирибонуклеїнова кислота) довжиною 160 мм укладається в об'ємі всього 0,5´10 мкм ) , ядерна мембрана руйнується і Х. вишиковуються на екваторі клітки. У цей період вони найбільш доступні для спостереження і вивчення їх морфології. Основна структурна одиниця метафазних Х., так само як і інтерфазних, — нитка ДНП діаметром 100—200, укладена в щільну спіраль. Деякі автори виявляють, що нитки діаметром 100—200 утворюють структури другого рівня укладання — нитки діаметром близько 2000, які і формують тіло метафазной Х. Каждая метафазная Х. складається з хроматид ( мал. 3 , 1 ), що утворилися в результаті реплікації вихідної інтерфазной Х. Використання мічених і модифікованих попередників ДНК(дезоксирибонуклеїнова кислота) дозволило чітко розрізняти в Х., що знаходиться в метафазі мітоза, диференціально забарвлені хроматиди, завдяки чому було встановлено, що при реплікації Х. незрідка відбувається обмін ділянками між сестринськими хроматидами (кросинговер ) . В класичній цитології надавалося велике значення матриксу метафазной Х., його вважали обов'язковим компонентом, в який занурені хромонеми, що спіралізують. Сучасні цитологи розглядають матрикс метафазних Х. як залишковий матеріал ядерця, що руйнується; часто він зовсім не виявляється.
Формування статевих кліток у тварин і рослин супроводиться особливим типом їх ділення — мейозом, і мейотічеськие Х. мають ряд особливостей в порівнянні з мітотичними. Перш за все, при мейозі дочірні клітки отримують удвічі зменшене число Х. (при мітозе воно зберігається однаковим), що досягається завдяки кон'югації гомологічних Х. у профазі мейозу і двома послідовними діленнями клітки при одній реплікації ДНК(дезоксирибонуклеїнова кислота) (детальніше за див.(дивися) Мейоз ) . Крім того в мейотічеськие Х. наголошуються тимчасова перерва профази мейозу і повернення їх до інтерфазному стану, коли Х. починають активно синтезувати РНК(рибонуклеїнова кислота). У цьому періоді у більшості вивчених тваринних організмів спостерігаються Х. типа лампових щіток ( мал. 4 ). Нарешті, Х. у метафазі мейозу відрізняються щільнішою упаковкою.
Не дивлячись на величезне число досліджень, присвячених Х., вивчення їх структурної і функціональній організації продовжує залишатися одним з найактуальніших напрямів сучасній біології. Х. виконують в клітці складні функції і мають вельми складну організацію, що важко піддається вивченню. Величезні успіхи в розумінні молекулярних основ будови Х. досягнуто в 60—70-і рр. 20 ст завдяки розвитку молекулярної генетики . Ці успіхи блискуче підтвердили основні положення хромосомній теорії спадковості, поглибивши і розвинувши їх.
Літ.: Вільсон Е., Клітка і її роль в розвитку і спадковості, пер.(переведення) з англ.(англійський), т. 1—2, М. — Л., 1936—40; Кольцов Н. До., Організація клітки, М. — Л., 1936; Прокофьева-Бельговськая А. А., Будова хромосоми, в кн.: Іонізуючі випромінювання і спадковість, М., 1960 (Підсумки науки. Біологічні науки, ст 3); Кикнадзе І. І., Функціональна організація хромосом Л., 1972; Де Робертіс Е., Новінський Ст, Саес Ф., Біологія клітки, пер.(переведення) з англ.(англійський), М., 1973; Левітський Р. А., Цитологія рослин. Ізбр. праці, М., 1976; Darlington С. D., Recent advances in cytology, 2 ed., L., 1937; Geitler L., Chromosomenbau, B., 1938 (Protoplasma-monographien, Bd 14); Ris Н., Kubai D. F., Chromosome structure, «Annual Review of Genetics», 1970, v. 4, р. 236—94; Handbook of molecular cytology, ed. by Lima-de-faria A., Amst. — L., 1969; Chromosome structure and function, N. Y., 1974.