Хромосомна теорія спадковості, теорія, згідно якої хромосоми, укладені в ядрі клітки, є носіями генів і є матеріальною основою спадковості, тобто спадкоємність властивостей організмів у ряді поколінь визначається спадкоємністю їх хромосом. Х. т.з. виникла на початку 20 ст на основі клітинній теорії і використання для вивчення спадкових властивостей організмів гибрідологичеського аналізу .
В 1902 У. Сеттон в США, що звернув увагу на паралелізм в поведінці хромосом і Менделеві т.з. «спадкових чинників», і Т. Бовері в Германії висунули хромосомну гіпотезу спадковості, згідно якої спадкові чинники (назва згодом генами) Менделя локалізовані в хромосомах. Перші підтвердження цієї гіпотези були отримані при вивченні генетичного механізму визначення підлоги у тварин, коли було з'ясовано, що в основі цього механізму лежить розподіл статевих хромосом серед нащадків. Подальше обгрунтування Х. т.з. належить американському генетикові Т. Х. Моргану, який відмітив, що передача деяких генів (наприклад, гена, обумовлюючого белоглазіє у самок дрозофіли при схрещуванні з червоноокими самцями) пов'язана з передачею статевої Х-хромосоми, тобто що успадковуються ознаки, зчеплені з підлогою (у людини відомо декілька десятків таких ознак, у тому числі деякі спадкові дефекти — дальтонізм, гемофілія і ін.).
Доказ Х. т.з. було отримано в 1913 американським генетиком До. Бріджесом, що відкрив нерозходження хромосом в процесі мейозу у самок дрозофіли і що відзначив, що порушення в розподілі статевих хромосом супроводиться змінами в спадкоємстві ознак, зчеплених з підлогою.
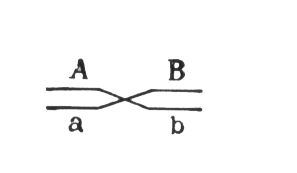
З розвитком Х. т.з. було встановлено, що гени, розташовані в одній хромосомі, складають одну групу зчеплення (див. Зчеплення генів ) і повинні успадковуватися спільно; число груп зчеплення дорівнює числу пар хромосом, постійному для кожного виду організмів (див. Каріотип ) ; ознаки, залежні від зчеплених генів, також успадковуються спільно. Внаслідок цього закон незалежного комбінування ознак (див. Менделя закони ) повинен мати обмежене вживання; незалежно повинні успадковуватися ознаки, гени яких розташовані в різних (негомологічних) хромосомах. Явище неповного зчеплення генів (коли поряд з батьківськими поєднаннями ознак в потомстві від схрещувань виявляються і нові рекомбінантниє, їх поєднання) було детально досліджено Морганом і його співробітниками (А. Р. Стертевантом і ін.) і послужило обгрунтуванням лінійного розташування генів в хромосомах. Морган передбачив, що зчеплені гени гомологічних хромосом, що знаходяться у батьків в поєднаннях і, в мейозі в гетерозиготної форми ® можуть мінятися місцями, внаслідок чого поряд з гаметамі АВ і ab утворюються гамети Ab і аВ. Подібні перекомбінациі відбуваються завдяки розривам гомологічних хромосом на ділянці між генами і подальшому з'єднанню розірваних кінців в новому поєднанні: Реальність цього процесу, названого перехрещенням хромосом, або кросинговером, була доведена в 1933 йому, ученим К. Штерномв дослідах з дрозофілой і американськими ученими Х. Крейтономі Б. Мак-Клінток — з кукурудзою. Чим далі друг від друга розташовані зчеплені гени, тим більше вірогідність кросинговера між ними. Залежність частоти кросинговера від відстаней між зчепленими генами була використана для побудови генетичних карт хромосом . В 30-х рр. 20 ст Ф. Добржанський показав, що порядок розміщення генів на генетичних і цитологичеських картах хромосом збігається.
Згідно з представленнями школи Моргана, гени є дискретними і далі неділимими носіями спадкової інформації. Проте відкриття в 1925 радянськими ученими Г. А. Надсоном і Г. С. Філіпповим, а в 1927 американським вченим Р. Меллером впливу рентгенівських променів на виникнення спадкових змін (мутацій ) в дрозофіли, а також вживання рентгенівських променів для прискорення мутаційного процесу в дрозофіли дозволили радянським ученим А. С. Серебровському, Н. П. Дубініну і ін. сформулювати в 1928—30 уявлень про подільність гена на дрібніші одиниці, розташовані в лінійній послідовності і здібні до мутаційних змін. У 1957 цих вистав були доведені роботою американського ученого С. Бензера з бактеріофагом Т4. Використання рентгенівських променів для стимулювання хромосомних перебудов дозволило Н. П. Дубініну і Б. Н. Сидорову виявити в 1934 ефект положення гена (відкритий в 1925 Стертевантом), тобто залежність прояву гена від місця розташування його на хромосомі. Виникло уявлення про єдність дискретності і безперервності в будові хромосоми.
Х. т.з. розвивається у напрямі поглиблення знань про універсальних носіїв спадкової інформації — молекули дезоксирибонуклеїнової кислоти (ДНК). Встановлено, що безперервна послідовність пурінових і пірімідінових підстав уздовж ланцюга ДНК(дезоксирибонуклеїнова кислота) утворює гени, міжгенні інтервали, знаки почала і кінця прочитування інформації в межах гена; визначає спадковий характер синтезу специфічних білків клітки і, отже, спадковий характер обміну речовин. ДНК(дезоксирибонуклеїнова кислота) складає матеріальну основу групи зчеплення у бактерій і багатьох вірусів (в деяких вірусів носієм спадкової інформації є рибонуклеїнова кислота ) ; молекули ДНК(дезоксирибонуклеїнова кислота), що входить до складу мітохондрій, пластид і ін. органоїдів клітки, служать матеріальними носіями цитоплазматичної спадковості.
Х. т. н., пояснюючи закономірності спадкоємства ознак у тварин і рослинних організмів, грає важливу роль в з.-х.(сільськогосподарський) науці і практиці. Вона озброює селекціонерів методами виведення порід тварин і сортів рослин із заданими властивостями. Деякі положення Х. т.з. дозволяють раціональніше вести з.-х.(сільськогосподарський) виробництво. Так, явище зчепленого з підлогою спадкоємства ряду ознак в з.-х.(сільськогосподарський) тварин дозволило до винаходу методів штучного регулювання підлоги в тутового шовкопряда вибраковувати кокони менш продуктивної підлоги, до розробки способу розділення курчат по підлозі дослідженням клоаки — відбраковувати півники і т.п. Найважливіше значення для підвищення врожайності багато з.-х.(сільськогосподарський) культур має використання поліплоїдії . На знанні закономірностей хромосомних перебудов грунтується вивчення спадкових захворювань людини.
Літ.: Морган Т. Р., Структурні основи спадковості, пер.(переведення) з англ.(англійський), М-кодом.—П., 1924; його ж, Вибрані роботи по генетиці, пер, з англ.(англійський), М-кодом.—Л., 1937; Актуальні питання сучасної генетики, М., 1966; Лобашев М. Е., Генетика, 2 видавництва, Л., 1967; Класики радянської генетики. [Сб. ст.], Л., 1968.