Генетичні карти хромосом , схеми відносного розташування зчеплених між собою спадкових чинників — генів . Р. до. х. відображують реально існуючий лінійний порядок розміщення генів в хромосомах (див. Цитологичеськие карти хромосом ) і важливі як в теоретичних дослідженнях, так і при проведенні селекційної роботи, т.к. позволяют свідомо підбирати пари ознак при схрещуваннях, а також передбачати особливості спадкоємства і прояву різних ознак в організмів, що вивчаються. Маючи Р. до. х., можна по спадкоємству «сигнального» гена, тісно зчепленого з тим, що вивчається, контролювати передачу потомству генів, що обумовлюють розвиток важко аналізованих ознак; наприклад, ген, що визначає ендосперм в кукурудзи і що знаходиться в 9-ій хромосомі, зчеплений з геном, що визначає знижену життєздатність рослини. Багаточисельні факти відсутності (всупереч Менделя законам ) незалежного розподілу ознак в гібридів другого покоління були пояснені хромосомною теорією спадковості . Гени, розташовані в одній хромосомі, в більшості випадків успадковуються спільно і утворюють одну групу зчеплення, кількість яких, т. о., відповідає в кожного організму гаплоїдному числу хромосом (див. Гаплоїд ). Американський генетик Т. Х. Морган показав, проте, що зчеплення генів, розташованих в одній хромосомі, в диплоїдних організмів (див. Діплоїд ) не абсолютне; у деяких випадках перед утворенням статевих кліток між однотипними, або гомологічними, хромосомами відбувається обмін відповідними ділянками; цей процес носить назва перехрещення, або кросинговера . Обмін ділянками хромосом (з генами, що знаходяться в них) відбувається з різною вірогідністю, залежною від відстані між ними (чим далі один від одного гени, тим вище вірогідність кросинговера і отже, рекомбінації). Генетичний аналіз дозволяє виявити перехрещення лише при відмінності гомологічних хромосом по складу генів, що при кросинговері приводить до появи нових генних комбінацій. Звичайна відстань між генами на Р. до. х. виражають як % кросинговера (відношення числа особин мутантів, що відрізняються від батьків іншим поєднанням генів, до загальної кількості вивчених особин); одиниця цієї відстані — морганіда — відповідає частоті кросинговера у 1%.
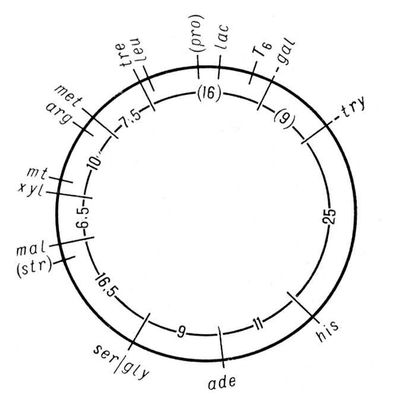
Р. до. х. складають для кожної пари гомологічних хромосом. Групи зчеплення нумерують послідовно, у міру їх виявлення. Окрім номера групи зчеплення, вказують повні або скорочені назви генів мутантів, їх відстані в морганідах від одного з кінців хромосоми, прийнятого за нульову крапку, а також місце центромери . Скласти Р.. х. можна лише для об'єктів, в які вивчено велике число генів мутантів. Наприклад, в дрозофіли ідентифіковано понад 500 генів, локалізованих в її 4 групах зчеплення, в кукурудзи — близько 400 генів, розподілених в 10 групах зчеплення ( мал. 1 ). В менш вивчених об'єктів число виявлених груп зчеплення менше гаплоїдного числа хромосом. Так, у будинкової миші виявлено близько 200 генів, створюючих 15 груп зчеплення (насправді їх 20); у курей вивчено доки всього 8 з 39. В людини з очікуваних 23 груп зчеплення (23 пари хромосом) ідентифіковано лише 10, причому в кожній групі відоме невелике число генів; найбільш детальні карти складені для статевих хромосом. У бактерій, які є гаплоїднимі організмами, є одна, найчастіше безперервна, кільцева хромосома і всі гени утворюють одну групу зчеплення ( мал. 2 ). При перенесенні генетичного матеріалу з клітки-донора в клітку-реципієнта, наприклад при кон'югації, кільцева хромосома розривається і лінійна структура, що утворюється, переноситься з однієї бактерійної клітки в іншу (в кишкової палички протягом 110—120 мін ). Штучно перериваючи процес кон'югації, можна по виниклих типах рекомбінантов встановити, які гени встигли перейти в клітку-реципієнта. У цьому полягає один з методів побудови Р. до. х. бактерій, детально розроблених в ряду видів. Ще більш деталізовані Р. до. х. деяких бактеріофагів . Див. також Генетика, Мутація .
Літ.: Лобашев М. Е., Генетика, 2 видавництва, Л., 1967; Медведев Н. Н., Практична генетика, 2 видавництва, М., 1968; Актуальні питання сучасної генетики. Сб. ст., М., 1966; Жакоб Ф., Вольман Е., Пів і генетика бактерій, пер.(переведення) з англ.(англійський), М., 1962; Бензер С., Тонка структура гена, в збірці: Молекулярна генетика, пер.(переведення) з англ.(англійський), М., 1963; Хейс В., Генетика бактерій і бактеріофагів, пер.(переведення) з англ.(англійський), М., 1965; Рейвін А. В., Еволюція генетики, пер.(переведення) з англ.(англійський), М., 1967; Мюнтцинг А., Генетика, пер.(переведення) з англ.(англійський), 2 видавництва, М., 1967: Уотсон Дж., Молекулярна біологія гена, пер.(переведення) з англ.(англійський), М., 1967.