Мейоз (від греч.(грецький) méiosis — зменшення), редукційне ділення, ділення дозрівання, спосіб ділення клітин, в результаті якого відбувається зменшення (редукція) числа хромосом в два рази і одну диплоїдну клітку (що містить два набори хромосом) після двох швидко наступних один за одним ділень дає початок 4 гаплоїдним (що містить по одному набору хромосом). Відновлення диплоїдного числа хромосом відбувається в результаті запліднення. М. — обов'язкова ланка статевого процесу і умова формування статевих кліток (гамет ). Біологічне значення М. полягає в підтримці постійності каріотипу у ряді поколінь організмів даного вигляду і забезпеченні можливості рекомбінації хромосом і генів при статевому процесі. М. — один з ключових механізмів спадковості і спадковою мінливості . Поведінка хромосом при М. забезпечує виконання основних законів спадковості (див. Менделя закони ).
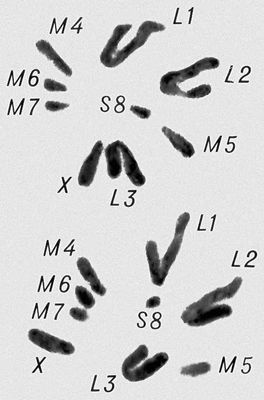
Залежно від місця М. в життєвому циклі організмів розрізняють 3 типи М. Гаметний, або термінальний, М. (у всіх багатоклітинних тварин і ряду нижчих рослин), відбувається в статевих органах і приводить до утворення гамет. Зіготний, або початковий, М. (в багатьох грибів і водоростей), відбувається в зіготе відразу після запліднення і приводить до освіти гаплоїдного міцелія або талому, а потім спор і гамет. Споровий, або проміжний, М. (у вищих рослин), має місце напередодні цвітіння і приводить до освіти гаплоїдного гаметофіта, в якому пізніше утворюються гамети. В простих (Protozoa) зустрічаються всі 3 типи М. Перед М. відбувається подвоєння кількості ДНК(дезоксирибонуклеїнова кислота) в клітці. В ході двох ділень М. ( мал. ) ДНК(дезоксирибонуклеїнова кислота) ділиться порівну між 4 клітками. В результаті першого (редукційного) ділення М. пари гомологічних хромосом відокремлюються і члени пар розходяться в 2 клітки (редукція числа хромосом). Кожна хромосома зберігає дві подовжні половини — хроматиди . В результаті другого (екваційного) ділення хроматиди розходяться в різні клітки і кожна з 4 сестринських кліток отримує по одній хроматиді. Т. о., перше ділення М. принципово відрізняється від мітоза, а друге — це мітоз в клітках з гаплоїдним числом хромосом. Під час М., перед редукцією числа хромосом, відбувається обмін ділянками гомологічних хромосом — кросинговер, що приводить до перерозподілу алельних генів (див. Аллелі ). М. триває багато довше за мітоза: наприклад, в пшениці він продовжується 24 години, в лілії — 9—12 діб, у миші — 11—14 діб, у людини — 24. В ряду тварин і людини під час утворення жіночих статевих кліток (див. Оогенез ) М. зупиняється на строк до декількох років і завершується лише під час запліднення.
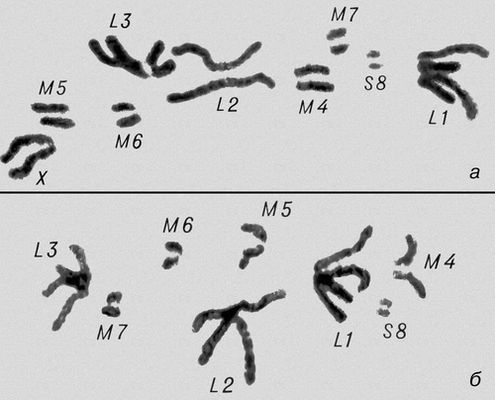
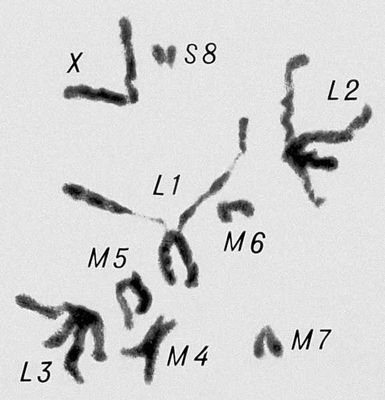
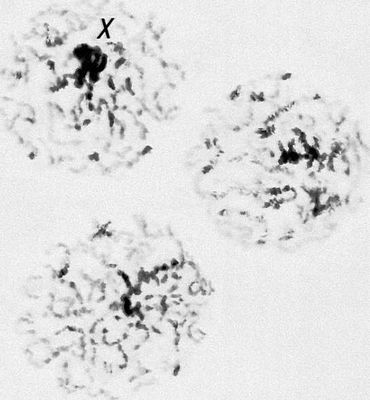
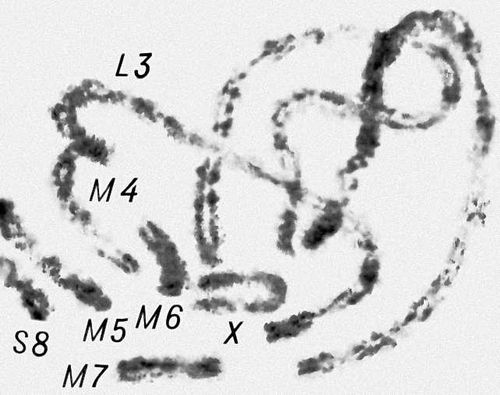
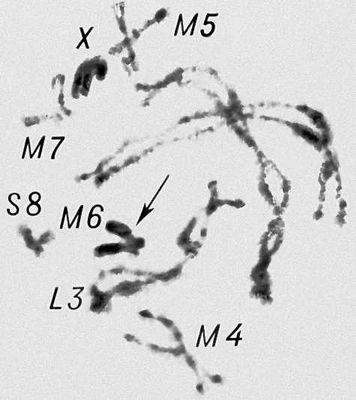
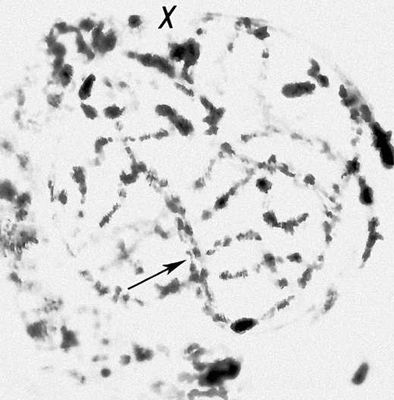
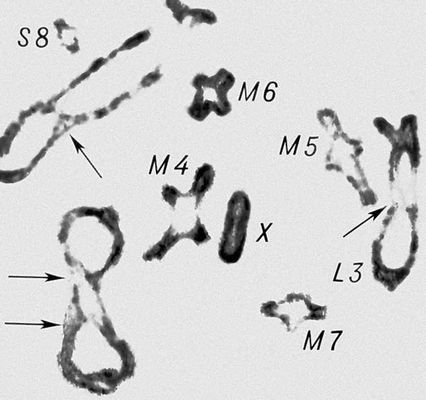
Перша фаза М. — профаза I, найбільш складна і тривала (у людини 22,5, в лілії 8—10 діб), підрозділяється на 5 стадій. Лептотена — стадія тонких ниток, коли хромосоми слабо спіралізують і найбільш довгі, видно потовщення — хромомери. Зіготена — стадія почала попарного, пліч-о-пліч з'єднання (синапсису, кон'югації) гомологічних хромосом; при цьому гомологічні хромомери взаємно притягуються і вишиковуються строго один проти одного. Пахитена — стадія товстих ниток; гомологічні хромосоми стабільно сполучені в пари — біваленти, число яких дорівнює гаплоїдному числу хромосом; під електронним мікроскопом видно складна ультраструктура в місці контакту двох гомологічних хромосом усередині бівалента: т.з. синаптонемальний комплекс, який починає формуватися ще в зіготене; у кожній хромосомі бівалента виявляються 2 хроматиди; т. о., бівалент (тетрада, по старій термінології) складається з 4 гомологічних хроматид; на цій стадії відбувається кросинговер, що здійснюється на молекулярному рівні; цитологичеськие наслідки його виявляються на наступній стадії. Діплотена — стадія ниток, що роздвоїлися; гомологічні хромосоми починають відштовхуватися один від одного, але виявляються зв'язаними, зазвичай в 2—3 крапках на бівалент, де видно хіазми (перехрещення хроматид) — цитологичеськоє прояв кросинговера. Діакенез — стадія відштовхування гомологічних хромосом, які як і раніше сполучені в біваленти хіазмами, що переміщаються на кінці хромосом (терміналізація); хромосоми максимально короткі і товсті (за рахунок тієї, що спіралізує) і утворюють характерні фігури: хрести, кільця і ін. Наступна фаза М. — метафаза I, під час якої хіазми ще зберігаються; біваленти вишиковуються в середній частині веретена ділення клітки, орієнтуючись центромерами гомологічних хромосом до протилежних полюсів веретена. У анафазе I гомологічні хромосоми за допомогою ниток веретена розходяться до полюсів; при цьому кожна хромосома пари може відійти до будь-якого з двох полюсів, незалежно від розбіжності хромосом ін. пар. Тому число можливих поєднань при розбіжності хромосом рівне 2 n , де n — число пар хромосом. На відміну від анафази мітоза, центромери хромосом не розщеплюються і продовжують скріпляти 2 хроматиди в хромосомі, що відходить до полюса. У телофазі I в кожного полюса починається деспіралізация хромосом і формування дочірніх ядер і кліток. Далі слідує коротка інтерфаза без редуплікації ДНК(дезоксирибонуклеїнова кислота) — інтеркінез, і починається друге ділення М. Профаза II, метафаза II, анафаза II і телофаза II проходят швидко; при цьому в кінці метафази II розщеплюються центромери, і в анафазе II розходяться до полюсів хроматиди кожної хромосоми. Ета класична схема М. має виключення. Наприклад, в рослин роду ожіка (Luzula) і комах сімейства кокцид (Coccidae) в першому діленні М. розходяться хроматиди, а в другому — гомологічні хромосоми, проте і в цих випадках в результаті М. відбувається редукція числа хромосом. Відмінності між сперматогенезом і оогенезом у тварин і утворенням мікроспор і мегаспор в рослин не відбиваються на поведінці хромосом в ході М., хоча розміри і долі сестринських кліток виявляються різними. Відомі аномалії М. В міжвидових гібридів всі хромосоми а в анеуплоїдов (див. Анеуплоїдія ) непарні хромосоми не здатні кон'югировать і залишаються у вигляді унівалентов; в автополіплоїдов (див. Автополіплоїдія ) утворюються об'єднання більш ніж з 2 хромосом — т.з. мультиваленти. У кожному з цих випадків неможлива правильна редукція числа хромосом в анафазе I; гамети, що утворюються (з незбалансованими наборами хромосом) або самі нежиттєздатні, або дають нежиттєздатне або потворне потомство (див. Хромосомні хвороби ). Відсутність хіазм (ахизматія) зазвичай приводить до тих же результатів, проте у самців деяких видів мух, у тому числі в дрозофіли, хіазми завжди відсутні, хоча гамети утворюються нормальні. Причини переходу кліток від ділення шляхом мітоза к М. у життєвому циклі кожного організму, а також молекулярні механізми кон'югації гомологічних хромосом і кросинговера досліджуються.
Літ.: Соколів І. І., Цитологичеськие основи статевого розмноження багатоклітинних тварин, в кн.: Керівництво по цитології, т. 2, М. — Л., 1966, с. 390—460; Райків І. Б., Карпология простих, Л., 1967; Богданов Ю. Ф., Гомологічна кон'югація хромосом, в кн.: Успіхи сучасної генетики, т. 3, М. 1971, с. 134—61; Swanson С. P., Cytology and cytogenetics, Englewood Cliffs, 1957; Rhoades М. М., Meiosis, в кн.: The cell. Biochemistry. Physiology. Morphology, v. 3, N. Y., 1961, р. 1—75; John B., Lewis K. R., The meiotic system, W. — N. Y., 1965; (Protoplasmatologia, Bd 6, fasc. 1); Westergaard М., Wettstein D. von, The Synaptinemal complex, в кн.: Annual Review of Genetics, v. 6, 1972, р. 71—110.