Мінливість (біологічна), різноманітність ознак і властивостей у особин і груп особин будь-якої міри спорідненості. І. властива всім живим організмам, тому в природі відсутні особини, ідентичні по всіх ознаках і властивостях. Термін «І.» уживається також для позначення здатності живих організмів відповідати морфофізіологичеськимі змінами на зовнішні дії і для характеристики перетворень форм живих організмів в процесі їх еволюції. І. можна класифікувати залежно від причин, природи і характеру змін, а також цілей і методів дослідження. Розрізняють І. спадкову (генотипічну) і неспадкову (паратипічну); індивідуальну і групову; переривисту (дискретну) і безперервну; якісну і кількісну; незалежну І. різних ознак і корелятивну (співвідносну); направлену (визначену, по Ч. Дарвіну) і ненапрямлену (невизначену, по Ч. Дарвіну); адаптивну (пристосовну) і неадаптивну. При вирішенні загальних проблем біології і особливо еволюції найбільш істотний підрозділ І., з одного боку, на спадкову і неспадкову, а з іншої — на індивідуальну і групову. Всі категорії І. можуть зустрічатися в спадковій і неспадковій, груповій і індивідуальній І.
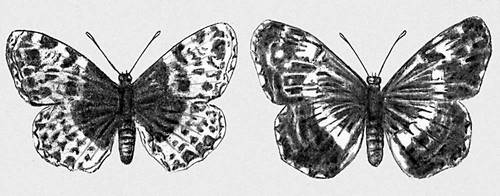
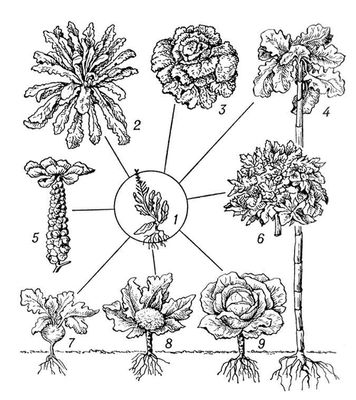
Спадкова І. обумовлена виникненням різних типів мутацій і їх комбінацій в подальших схрещуваннях. У кожній досить тривалий (у ряді поколінь) існуючій сукупності особин спонтанно і ненапрямлено виникають різні мутації, які надалі комбінуються більш менш випадково з різними вже наявними в сукупності спадковими властивостями. І., обумовлену виникненням мутацій, називають мутаційною, а обумовлену подальшим перекомбінірованієм генів в результаті схрещування — комбінаційною. На спадковій І. заснована вся різноманітність індивідуальних відмінностей, які включають: а) як різкі якісні відмінності, не зв'язані один з одним перехідними формами, так і чисто кількісні відмінності, створюючі безперервні ряди, в яких близькі члени ряду можуть відрізнятися один від одного скільки завгодно мало; б) як зміни окремих ознак і властивостей (незалежна І.), так і взаємозв'язані зміни ряду ознак (корелятивна І.); у) як зміни, що мають пристосовне значення (адаптивна І., мал. 1 ), так і зміни «байдужі» або навіть знижуючі життєздатність їх носіїв (неадаптивна І.). Всі ці типи спадкових змін складають матеріал еволюційного процесу (див. Мікроеволюція ). У індивідуальному розвитку організму прояв спадкових ознак і властивостей завжди визначається не лише основними, відповідальними за дані ознаки і властивості генами, але і їх взаємодією з багатьма іншими генами, складовими генотип особини, а також умовами зовнішнього середовища, в якому протікає розвиток організму ( мал. 2 і 3 ).
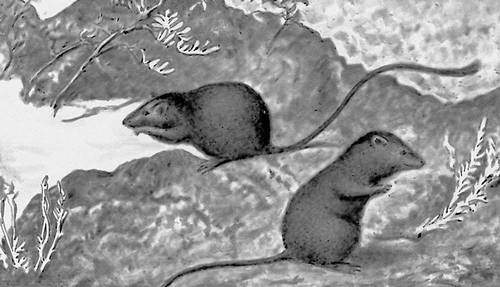
В понятіє неспадковою І. входять ті зміни ознак і властивостей, які у особин або певних груп особин викликаються дією зовнішніх чинників (живлення, температура, світло, вологість і т. д.). Такі неспадкові ознаки (модифікації ) в їх конкретному прояві у кожної особини не передаються по спадку, вони розвиваються у особин подальших поколінь лише за наявності умов, в яких вони виникли. Така І. називається також модифікаційною ( мал. 4 ). Наприклад, забарвлення багатьох комах при низькій температурі темніє, при високій — яснішає; проте їх потомство буде забарвлено незалежно від забарвлення батьків відповідно до температури, при якій воно само розвивалося (див. Морфози, Фенокопія ). Існує ще одна форма неспадковою І. — так звані тривалі модифікації, що часто зустрічаються в одноклітинних організмів, але зрідка спостережувані і в багатоклітинних. Вони виникають під впливом зовнішніх дій (наприклад, температурних або хімічних) і виражаються в якісних або кількісних відхиленнях від вихідної форми, зазвичай поступово затухаючих при подальшому розмноженні. Вони засновані, мабуть, на змінах відносно стабільних цитоплазматичних структур.
Між неспадковою і спадковою І. існує тісний зв'язок. Неспадкових (в буквальному розумінні) ознак і властивостей немає, оскільки неспадкові зміни є віддзеркаленням спадково обумовленої здатності організмів відповідати певними змінами ознак і властивостей на дії чинників зовнішнього середовища. При цьому межі неспадкових змін визначаються нормою реакції генотипу на умови середовища.
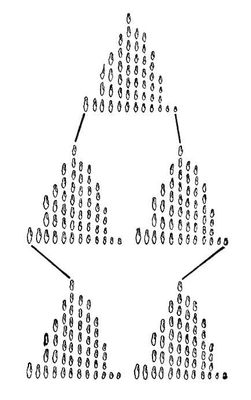
Спадкову і неспадкову І. вивчають як усередині окремих совокупностей живих організмів, коли досліджують відмінності ознак окремих особин (індивідуальна І.), так і при порівнянні між собою різних совокупностей особин (групова І.); у основі будь-яких міжгрупових відмінностей також лежить індивідуальна І. Даже в межах близькоспоріднених груп немає абсолютно ідентичних особин, які не розрізнялися б по мірі вираженості яких-небудь спадкових або неспадкових ознак і властивостей. Зважаючи на складність організації живих систем, навіть в генотіпічеськи ідентичних (наприклад, однояйцеві близнята) особин, що розвиваються в практично однакових умовах, завжди можна виявити хоч би незначні морфофізіологичеськие відмінності, пов'язані з неминучими флуктуаціями умов середовища і процесів індивідуального розвитку. Групова І. включає відмінності між совокупностямі будь-яких рангів — від відмінностей між невеликими групами особин в межах популяції до відмінностей між царствами живої природи (тварини — рослини). По суті, вся систематика організмів побудована на порівняльному аналізі групової І. Для вивчення пускових механізмів еволюційного процесу особливе значення мають різні форми внутрівидової групової І. (див. Відообразованіє ). Більшість видів розпадається на підвиди або географічні раси. В разі повної ізоляції географічних форм вони можуть різко розрізнятися поодинці або декільком ознакам. Популяції, що населяють обширні території і не розділені різкими ізолюючими бар'єрами, можуть (завдяки перемішуванню і схрещуванню) поступово переходити один в одного, утворюючи кількісні градієнти по тих або інших ознаках (клінальная І.). Географічна, у тому числі і клінальная, І. у природних умовах — результат дії ізоляції, природного відбору і ін. чинників еволюції, що приводять до розділення вихідної групи особин в ході історичного формування вигляду на дві або декілька груп, що розрізняються по чисельних співвідношеннях генотипів ( мал. 5 ). В деяких випадках відмінності між групами особин в межах вигляду не пов'язані з відмінностями їх генотипічного складу, а обумовлюються модифікаційною І. (різними реакціями схожих генотипів на різні зовнішні умови). Так звана сезонна І. обумовлена впливом на розвиток відповідних поколінь різних погодних умов (наприклад, у деяких комах і трав'янистих рослин, що дають два покоління в рік, весняні і осінні популяції розрізняються рядом ознак) ( мал. 6 ). Інколи сезонні форми можуть бути результатом відбору різних генотипів (наприклад, рано- і поздноцветущие форми трав на сінокісних лугах: протягом багатьох поколінь усувалися особини, квітучі влітку, під час сінокосу). Великий інтерес представляє екологічна І. — відмінності між групами особин одного вигляду, що зростаючими або живуть в різних місцях (піднесеності і низовині, заболочені і сухі ділянки і т. д.). Часто такі форми називаються екотипами . Виникнення екотипів також може бути результатом як модифікаційних змін, так і відбору генотипів, краще пристосованих до місцевих умов. Спадковою І. обумовлені різні форми внутрішньопопуляції поліморфізму . У деяких популяціях спостерігається співіснування два або більш за ясно помітні форми (наприклад, у двоточкового сонечка майже у всіх популяціях зустрічаються чорна форма з червоними плямами і червона форма з чорними плямами). У основі цього явища можуть лежати різні еволюційні механізми: неоднакова пристосованість форм, що співіснують, до умов різних сезонів року, підвищена життєздатність гетерозигот, в потомстві яких постійно вищепляются обидві гомозиготні форми або інші, ще недостатньо вивчені механізми. Таким чином, і групова, і індивідуальна І. включають зміни як спадкової, так і неспадкової природи.
Незалежною І. ознак протиставляють корелятивну І. — взаємозв'язана зміна різних ознак і властивостей: зв'язок між зростанням і вагою особин (позитивна кореляція) або темпом клітинного ділення і величиною кліток (негативна кореляція). Кореляції можуть бути обумовлені чисто генетичними причинами (плейотропія ) або взаїмозавісимостямі процесів становлення певних ознак і властивостей в індивідуальному розвитку особин (онтогенетичні кореляції), а також схожими реакціями різних ознак і властивостей на одні і ті ж зовнішні дії (фізіологічні кореляції). Нарешті, кореляції можуть відображати історію походження популяцій з суміші два або більш за форми, кожна з яких привносить не окремі ознаки, а комплекси взаємозв'язаних ознак і властивостей (історичні кореляції). Вивчення корелятивної І. має важливе значення в палеонтології (наприклад, при реконструкції вимерлих форм по окремих викопних залишках), у антропології (наприклад, при відновленні рис обличчя на основі вивчення черепа), в селекції і медицині.
Основні методи вивчення І. — порівняльно-описовий і біометричний (див. Біометрія ). Сукупність цих методів дозволяє досліджувати як паратипічну, так і генотипічну компоненти загальної фенотипічної І. Так, першу можна вивчати, порівнюючи генотіпічеськи ідентичні клони і чисті лінії, що розвиваються в різних умовах. Складніше виділити чисто генотипічну І. із загальної фенотипічної. Це можливо зробити на основі біометричного аналізу (див. Наслідуваність ). У медичній генетиці для тих же цілей використовується визначення відсотка конкордантності (збіги) тих або інших ознак у одно- і різнояєчних близнят.
Спадковість і І. живих організмів інколи протиставляють як «консервативне» і «прогресивне» почала. Насправді ж вони найтіснішим чином зв'язані. Відсутність повної стабільності генотипу обумовлює мутаційну і (в ході подальших схрещувань і розщеплювань) комбінаційну І., тобто в цілому — генотипічну І. Паратіпічеськая (неспадкова) І. — результат лише відносній стабільності генотипу при визначенні ним в онтогенезі норми реакції при розвитку ознак і властивостей особин. З цього виходить можливість експериментальних дій як на спадкову, так і на неспадкову І. Первую можна підсилити дією мутагенних чинників (випромінювання, температура, хімічні речовини). Розмах і напрям комбінаційної І. можна контролювати за допомогою штучного відбору . На неспадкову І. можна впливати, змінюючи умови середовища (живлення, світло, вологість і т. д.), в яких протікає розвиток організму.
Чітке уявлення про категорії і форми І. необхідно при побудові еволюційних схем і теорій, оскільки явища спадковості і І. лежать в основі еволюційного процесу, а також в практичній селекції рослин і тварин, при вивченні ряду проблем медичної географії і антропології популяції.
Літ.: Філіпченко Ю. А., Мінливість і методи її вивчення, 2 видавництва, Л., 1926; Четвериків С. С., Про деякі моменти еволюційного процесу з точки зору сучасної генетики, «Журнал експериментальної біології», 1926, т. 2 № 1; Іогансен Ст, Елементи точного вчення про мінливість і спадковості з основами варіаційної статистики, М. — Л., 1933; його ж, Про спадкоємство в популяціях і чистих лініях, М. — Л., 1935; Холден Дж., Чинники еволюції, пер.(переведення) з англ.(англійський), М. — Л., 1935; Дарвін Ч., Походження видів ..., Соч., т. 3, М., 1939; Шмальгаузен І. І., Організм, як ціле в індивідуальному і історичному розвитку, [2 видавництва], М. — Л., 1942; Астауров Би. Л., Мінливість, в кн.: Велика медична енциклопедія, т. 11, М., 1959; Вавілов Н. І., Закон гомологічних рядів в спадковій мінливості, Ізбр. проїзв.(твір), т. 1, Л., 1967, с. 7—61; його ж, вигляд як система Ліннєєвський, там же, с. 62—87; Лобашев М. Е., Генетика, 2 видавництва, Л., 1967; Майр Е., Зоологічний вигляд і еволюція, пер.(переведення) з англ.(англійський), М., 1968; Тімофєєв-Ресовський Н. Ст, Воронцов Н. Н., Я блоків А. Ст, Короткий нарис теорії еволюції, М., 1969; Fisher R., The genetical theory of natural selection, Oxf., 1930; Falconer D., Introduction to quantative genetics, Edinburgh — L., 1960.
Н. Ст Тімофєєв-Ресовський, Е. До. Гинтер, Н. Ст Глотов, Ст І. Іванов.
Мінливість у мікроорганізмів. У мікроорганізмів, як і в інших організмів, розрізняють неспадкову і спадкові І. Ізмененію можуть піддаватися будь-які морфологічні і фізіологічні ознаки: величина і форма мікроорганізмів, вигляд і забарвлення їх колоній, здатність засвоювати або синтезувати різні органічні речовини, хвороботворність і ін. Спадкова І. мікроорганізмів — результат мутацій, що виникають спонтанно або що викликаються фізичними або хімічними мутагенами (ультрафіолетові промені, іонізуюча радіація, етиленимін і ін.). У мутантів можуть різко посилюватися або знижуватися такі кількісні ознаки, як здібність до біосинтезу амінокислот, антибіотиків, ферментів, вітамінів і тому подібне Виникають так звані дефіцитні мутанти, здатні зростати лише при додаванні до середовища певних амінокислот, пуринів, пірімідінов і ін. Мікроорганізми розмножуються дуже швидко. Тому на них легко вивчати всі форми І., а також здійснювати штучний відбір корисних мутантів (див. Селекція ). Так, при безперервному культивуванні відповідних мікроорганізмів (проточні культури) в живильному середовищі, що містить, наприклад, антибіотик, фенол або сулему, легко можуть бути отримані форми, стійкі до даної речовини (адаптивна І.). Спостерігаються у мікроорганізмів і взаємозв'язані зміни (корелятивна І.). Так, виникнення у хвороботворних мікробів складчастих колоній супроводиться зниженням їх імуногенності. У мікроорганізмів, що мають дійсний статевий процес (деякі плісневі гриби, спорогенні дріжджі) можливе схрещування, що супроводиться перекомбінірованієм генів і здобуттям гібридів. В недосконалих грибів і бактерій, позбавлених дійсного статевого процесу, такі гібриди не можуть бути отримані.