Пів організмів, сукупність морфологічних і фізіологічних особливостей організму, що забезпечують статеве розмноження, суть якого зводиться зрештою до заплідненню . При цьому чоловічі і жіночі статеві клітки — гамети зливаються в зіготу, з якої розвивається новий організм. У зіготе об'єднуються 2 гаплоїдних (одинарних) набору хромосом материнської і батьківської гамет. У статевих клітках нового організму утворюються гаплоїдниє набори вже перекомбінірованних (див. Рекомбінація ) батьківських і материнських хромосом (в результаті обміну ділянками гомологічних батьківських хромосом — кросинговера — і випадкової їх розбіжності по дочернім кліткам під час мейозу ) . Тому в обох статей популяції постійно виникає безліч генетично різних особин, що створює сприятливі умови для природного відбору більш пристосованих форм. У цьому полягає основна перевага статевого розмноження перед безстатевою. Статеве розмноження переважає у тварин і вищих рослин; воно зустрічається і у багатьох мікроорганізмів (кон'югація у бактерій супроводиться частковим обміном спадковим матеріалом — нитками ДНК(дезоксирибонуклеїнова кислота)). Статевий процес в одноклітинних організмів не вимагає значної диференціація П. (одна і та ж клітка може бути і кліткою тіла, і половою). В багатоклітинних диплоїдних організмів виникли спеціальні гаплоїдниє статеві клітки: великі і малорухливі або нерухомі в жіночого П., дрібні і зазвичай рухливі — в чоловічого. У більшості рослин і лише у деяких тварин обидва типи гамет виробляються однією особиною (див. Гермафродитизм ) , у більшості тварин — різними особинами, які у зв'язку з цим строго розділяються відповідно на самок і самців. Окрім продукування кліток різного П., самці і самки розрізняються рядом морфологічних і фізіологічних ознак, а також статевою поведінкою, які забезпечують злиття статевих кліток.
Визначення підлоги. Всі організми, у тому числі і раздельнополиє, в генетичному відношенні бісексуальні (двостатеві) т.к. зиготы їх отримують генетичну інформацію, можливість розвивати ознаки чоловічого і жіночого П. В обох статей рослин і деяких гермафродитних тварин жіночі і чоловічі репродуктивні органи і статеві клітки розвиваються з генетично однакових кліток під впливом внутрішніх умов, що потенційно дає (по відношенню до окремих кліток їх можна розглядати як зовнішні). Механізм перемикання кліток на розвиток в одному випадку жіночих, в іншому чоловічих репродуктивних органів повністю не розкритий. У окремих випадках в раздельнополих видів потенційно бісексуальні зіготи розвиваються в самок або самців під впливом зовнішніх умов. Наприклад, у морського кільчастого черв'яка бонеллія личинка, поселяясь на хоботку самки, розвивається в самця, а на дні морить — в самку. В рослини Arisaema japonica з крупних бульб, багатих живильними речовинами, розвиваються рослини з жіночим квітками, а з дрібних бульб — з чоловічими. Визначення П. під впливом зовнішніх умов називається фенотипічним, або модифікаційним.
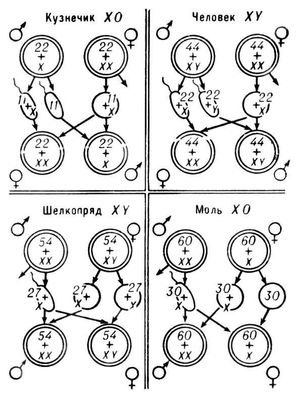
Ширше поширене генетичне визначення П. В цьому випадку зігота під час запліднення також дістає потенційні можливості для розвитку ознак обидва П. Однако під впливом генетичних чинників в одній половині зігот пересилює тенденція розвитку чоловічого П., а в іншій — жіночого. Спеціальний хромосомний механізм забезпечує передачу одній половині потомства генів жіночого П., а інший — генів чоловічого П. На початку 20 ст було встановлено, що у самців деяких видів комах в диплоїдних (з подвійним набором хромосом) клітках поряд з парами гомологічних хромосом є одна непарна хромосома. Самка ж має дві такі хромосоми. У самців комах ін. видів всі хромосоми парні, але в одній з пар вони морфологічно несхожі. Ці хромосоми, причетні до визначення П., назвали статевими хромосомами, а останні — аутосомами . Пізніше статеві хромосоми були виявлені в багатьох раздельнополих організмів. Статеву хромосому самця, що повторюється у самок, назвали Х-хромосомою, а не що повторюється — y-хромосомою. Поєднання статевих хромосом самця позначають формулою X0 або XY, а самки — XX. Самці з однією статевою хромосомою продукують в рівній кількості гамети з Х-хромосомою і гамети, позбавлені її, тобто з одним лише гаплоїдним набором аутосом (А); самки — гамети лише з Х-хромосомою. Після випадкового злиття чоловічих і жіночих гамет половина зігот, що утворилися, матиме дві Х-хромосоми (XX), а ін. половина — лише одну Х-хромосому. Перші стануть самками, другі — самцями ( мал. ).
Самці з різними статевими хромосомами продукують в рівній кількості гамети, що мають Х-хромосому, і гамети, що мають y-хромосому. Жіночі гамети цього вигляду генетично однакові — всі вони несуть по одній Х-хромосомі. В результаті половина яйцеклітин буде запліднена сперматозоїдами з y-хромосомою, а ін. половина — з Х-хромосомою. Перші зіготи, що мають структуру XY, розвинуться в особини чоловічого П., другі — з XX — в особини жіночого П. Самци з однією Х-хромосомою або з двома різними (XY) хромосомами мають гетерогаметний П., самки з ХХ-хромосомамі — гомогаметний П. У багатьох тварин, навпаки, самки мають гетерогаметний П. Іх статеві хромосоми позначають буквами Z і W або XY, а статеві хромосоми гомогаметних самців — ZZ або XX. У ссавців, нематод, молюсків, голкошкірих і у більшості членистоногих гетерогаметен чоловічий П. У комах і риб гетерогаметність спостерігається як в чоловічого, так і в жіночого П. Гетерогаметность жіночого П. властива птицям, плазунам і деяким земноводним.
Бісексуальні потенції, властиві зіготе, обумовлені генами, локалізованими в аутосомах і такими, що виявляються лише під контролем ін. генів — реалізаторів П. Іменно ці гени відкривають дорогу в одному випадку генам, сприяючим утворенню жіночого П., в іншому — генам, що обумовлюють розвиток чоловічого П. Прі генетичному визначенні П. за типом X0, XX реалізаторів жіночого П. локалізовано в Х-хромосомах, а чоловічого — в аутосомах. При поєднанні однієї дози реалізаторів жіночого П., локалізованих в одній Х-хромосомі, з диплоїдним набором реалізаторів чоловічого П., локалізованих в аутосомах, розвивається чоловічий П. І лише 2 дози реалізаторів жіночого П., локалізованої в 2 Х-хромосомах, пересилюють потенцію розвитку чоловічого П. і тим самим обумовлюють жіночий П. У людини полоопределяющую роль грає y-хромосома. У аномальних випадках вона поєднується з 2, 3 і навіть 4 Х-хромосомами при нормальному наборі аутосом. Хоча це і приводить до патологічних відхилень, проте всі особини з такими наборами хромосом бувають чоловічого П. Полоопределяющая роль y-хромосом відмічена в багатьох видів тварин, а серед рослин — в дрімоти лугової. В дрозофіли y-хромосома майже не містить генів, тобто спадково інертна; реалізатори жіночого П. локалізовані в Х-хромосомі, реалізатори чоловічого П. — в аутосомах. Розвиток П. контролюється відношенням Х-хромосом до набору аутосом (Х: А), умовно прийнятим у самки за одиницю (2Х:2А = 1): це відношення у самця дорівнює 0,5 (Х:2А = 0,5). Збільшення цього відношення (статевого індексу) зверху одиниці приводить до надмірного розвитку жіночих статевих ознак («надсамки»), зменшення ж нижче 0,5 сприяє появі самців з більш вираженими чоловічими ознаками («надсамці»). Особини із статевим індексом 0,67 і 0,75 мають проміжний розвиток ознак обидва П. і називають інтерсексамі. Явище інтерсексуальності демонструє бісексуальну потенцію спадкової інформації, передаваної всім нащадкам.
Механізм генетичного контролю над розвитком статевих ознак може бути внутрі- і міжклітинним. Внутріклітинне визначення П. не пов'язане з утворенням статевих гормонів (наприклад, у комах), і дія генів, що визначають П., обмежена клітками, в яких ці гени функціонують. При цьому в одному організмі можуть нормально розвиватися, не впливаючи один на одного, ділянки тіла з жіночими і чоловічими ознаками (див. Гинандроморфізм ) . При міжклітинному визначенні П., характерному для ссавців і птиць, під контролем генів виробляються статеві гормони, які, проникаючи у всі клітки організму, обумовлюють фенотипічний розвиток ознак відповідного П. Разлічают прогамне, сингамне і епігамне визначення П. Прогамноє визначення П. відбувається до запліднення яйця, наприклад диференціювання яйцеклітин на швидко і повільно зростаючі. Перші стають великими, і з них після запліднення розвиваються самки, другі відрізняються меншими розмірами і дають самців, хоча обидва види яйцеклітин генетично однакові. Сингамне визначення П. відбувається під час запліднення але на різних стадіях цього процесу. В деяких видів з чоловічою гетерогаметієй і фізіологічною поліспермієй (запліднення яйцеклітини декількома сперматозоїдами) П. визначається у момент злиття ядер статевих кліток (каріогамія ) . Якщо з ядром яйцеклітини зливається чоловіче ядро з y-хромосомою, розвинеться чоловіча особина, якщо з Х-хромосомою — жіноча. При жіночій гетерогаметії П. потомства залежить від того, яка із статевих хромосом потрапляє в ядро яйцеклітини під час мейозу. Якщо в ядрі виявиться z-хромосома, розвинеться особина чоловічого П., якщо w-хромосома — жіночого. Т. о., в даному випадку П. зіготи встановлюється до каріогамії. Епігамне визначення П. спостерігається в різностатевих видів з фенотипічним визначенням П., коли спрямованість розвитку у бік чоловічого або жіночого П. обумовлюється впливом зовнішніх умов після запліднення.
Залежність ознак від підлоги . Залежать від П. ознаки, обмежені і контрольовані їм. Ограниченные П. ознаки через статеву диференціацію можуть виявитися лише в одного з П. (продукція молока або яєць властива лише жіночому П.), хоча полімерні гени цих ознак локалізовані в аутосомах обидва П. Ознаки, контрольовані П., виявляються або в обох П. (з різною мірою вираженості), або (частіше) лише в одного з П. (потужніший розвиток рогів у баранів, бороди — у козлів), хоча обидва в рівній мірі містять в аутосомах гени цих ознак. Несхожий їх розвиток обумовлений значною відмінністю фізіологічних процесів в організмах різного П.
Гени, що детермінують ознаки, зчеплені з П., локалізовані як в парних, так і непарних статевих хромосомах і тому успадковуються інакше, ніж ознаки, обумовлені парними генами, локалізованими в аутосомах обидва П. Еслі гени локалізовані в непарній y-хромосомі гетерогаметного самця, то обумовлювані ними ознаки успадковуються лише синами, а при локалізації генів в хромосомі гетерогаметной самки — лише дочками. Успадковані т.ч. ознаки називаються голандрічеськимі. Цей тип спадкоємства виявлений в деяких видів риб і комах. В ін. видів тварин він з повною достовірністю не доведений. При локалізації генів в гомологічних Х- або Z- хромосомах обумовлені ними ознаки передаються зчеплений с П. за типом, що отримав назву спадкоємства навхрест, коли рецесивна ознака матері виявиться у синів, а домінантний — у дочок (Т. X. Морган), що зустрічається в багатьох видів тварин (наприклад, трибарвна кішок, смугастість забарвлення оперення і швидкість його росту у курей). Багато зчеплених с П. мутацій виявлено в дрозофіли і тутового шовкопряда.
Зчепленими с П. можуть бути і літали — гени, що обумовлюють смертельний результат при розвитку організму. Якщо гомогаметний батько гетерозіготен по літали, локалізованою в одній з гомологічних статевих хромосом (X або Z), то половина його гетерогаметних нащадків загине, отримавши деталь, згубній дії якої в генотипі не протиставить нормальний аллель . При гетерогаметії жіночого П. від леталей гине половина дочок, а при гетерогаметії чоловічого П. — половина синів. Інколи гени мутантів в Х- і Z- хромосомах лише частково знижують життєздатність потомства або викликають різні захворювання, що найчастіше виявляються в гетерогаметного П. У людини виявлено понад 50 зчеплених с П. мутацій, що приводять переважно до порушення нормальної життєдіяльності організму (див. Хромосомні хвороби ) .
Співвідношення полови . При фенотипічному визначенні П. воно залежить від кількості організмів, що розвиваються, які попадають під вплив зовнішніх чинників, що детермінують той або інший П. Прі генетичному визначенні П. співвідношення П. у більшості видів, як правило, дуже близько до 100♀: 100♂ (100 самок: 100 самців). Проте і при такому визначенні П. є відхилення. Так, в деяких видів ссавців з чоловічою гетерогаметієй статистично достовірно народжується на 1—2% більше нащадків чоловічого П.
Регуляція підлоги . Істотне зрушення співвідношення організмів у бік одного з П. має як теоретичне, так і практичне значення, т.к. одін з П. зазвичай продуктивніший. Методи регуляції П., зведені до 4 основних напрямів, застосовуються залежно від типа визначення П. і біологічних і господарських особливостей вигляду.
Фенотипічне перевизначення П. Еслі дія генів П. реалізується за допомогою гормонів, статеві ознаки змінюються при пересадці статевих органів одного П. іншому (див. Гонади ) або при введенні в організм гормонів протилежного П., а також деяких амінокислот. Міра фенотипічних змін П. залежить від особливостей вигляду і дози введеного препарату. Проте лише в окремих випадках (у деяких риб і земноводних) особини з фенотіпічеськи перевизначеним П. продукують гамети, протилежні до їх генотипічного П. В наступному поколінні, якщо дія гормонів припиняється, знову набирає чинності генетичний механізм визначення П.
Управління генетичним механізмом визначення П., або штучне поєднання в яйцеклітині статевих хромосом. Направлена зміна співвідношення П. досягнута в експериментах з тутовим шовкопрядом, в якого П. строго визначається поєднанням статевих хромосом (ZW — ♀ ; ZZ — ♂). Незапліднені яйця після прогрівання розвиваються партеногенетічеськи (див. Партеногенез ) за рахунок диплоїдного ядра, що не завершило редукційного ділення. Всі клітки партеногенетичного ембріона зберігають материнську структуру, зокрема і відносно статевих хромосом ZW, і, отже, розвиваються лише в самок (Би. Л. Астауров). Дією іонізуючих випромінювань і прогріванням удалося подавити в свіжовідкладеному заплідненому яйці жіноче ядро і перемкнути розвиток на чоловіче начало. Диплоїдне ядро чоловічої зіготи утворюється шляхом злиття двох чоловічих ядер і тому має структуру чоловічого П. ZZ. З таких зігот розвиваються гусениці завжди чоловічої статі (X. Хасимото; Б. Л. Астауров). Цими методами вперше в з.-х.(сільськогосподарський) вигляду шовкопряда вирішена проблема довільній регуляції П. У ссавців учені намагаються розділити по морфологічних і фізіологічних особливостях Х- і y-сперматозоїді з метою подальшого запліднення однією категорією сперматозоїдів. Проте цим способом доки не удалося достовірно змістити співвідношення П.
Раннє розпізнавання П. Іспользуєтся для сортування курчат, що вилупилися, на півники і курок по забарвленню оперення, зчепленому з П., а також для «надраннього» сортування по П. тутового шовкопряда. Під дією іонізуючого опромінення в шовкопряда пересаджена аутосома з домінантним геном, що обумовлює темне забарвлення яєць тутового шовкопряда, на статеву хромосому W-. Зчеплення хромосом стійко передається по спадку. Ті яйця, в які потрапляє W- хромосома з пересадженим домінантним геном, набувають темного кольору і розвиваються в самок, тоді як яйця чоловічого П., не отримавши домінантного гена, залишаються непігментованими. Фотоелектричні автомати з великою швидкістю розділяють різнозабарвлені яйця по П. Виведенниє в такий спосіб (В. А. Струнников і Л. М. Гуламова) мічені по П. породи шовкопряда знаходять практичне вживання в радянському шовківництві. У 60-х рр. 20 ст в дослідах англійських учених Р. Едуардса і Р. Гарднера зафіксовано народження потомства лише одного П. і у ссавців. У кроликів витягували з тіла матерів ранніх зародків, цитологичеським методом визначали їх П. і потім зародки небажаного П. вибраковували, а зародки потрібного П. повертали в матку. Близько 20% повернених зародків прижилося і розвивалося в крольчат передбаченого ученими П.
Зміна співвідношення П. може бути майже у всіх тварин з генетичним визначенням П. результатом загибелі половини зародків гетерогаметного П. під дією зчеплених с П. деталей. Проте для багатьох з.-х.(сільськогосподарський) тварин такий підхід до регуляції П. економічно не виправданий. Виняток становить тутовий шовкопряд. У СРСР радіаційним методом виведена (Ст А. Струнників) генетично особлива порода тутового шовкопряда, в якої в обох Z- хромосомах самців завжди є по одній негомологічній один одному літали (збалансовані літали). Якщо цих самців схрестити з самками звичайних порід, на стадії яйця одна половина самок загине від першої, а інша — від другої літали. З яєць чоловічого П. вилуплюються нормальні гусениці. Цей спосіб дозволяє в необмежених кількостях отримувати в тутового шовкопряда лише один продуктивніший чоловічий П.
Еволюція підлоги . Раздельнополость (бісексуальність), властива вже багатьом одноклітинним (водорості, прості), сталася від смешаннополості. Лише в деяких випадках (наприклад, при паразитизмі ) смешаннополость могла виникнути повторно з раздельнополості. Так, в паразитичних ракоподібних спостерігаються всі переходи від смешанно- до раздельнополості (наприклад, раздельнополиє види з добре розвиненими самками і карликовими самцями — виразне зрушення у бік гермафродитизму). Фенотипічне визначення П. древньо генетичного, т.к. на ранніх стадіях еволюції ще не існувало спеціального апарату статевих хромосом. Еволюції, що виникають на певних етапах, спеціальні статеві хромосоми (риби, земноводні) спочатку морфологічно невідмітні від аутосом, і про наявність їх можна судити лише по ознаках, зчепленим с П. Услід за морфологічними відмінностями між статевими хромосомами і аутосомами виникає диференціювання між Х- і Y- хромосомою, що робить усе більш рідкою кон'югацію між ними і утрудняє обмін їх ділянками при кросинговері (перехрещенні). Все це сприяє виконанню специфічної функції статевих хромосом — бути реалізаторами жіночого або чоловічого П. Полноє зникнення Y- хромосом робить генетичне визначення П. ще досконалішим: П. визначається рівновагою між числом аутосом і хромосом.
Літ.: Астауров Би. Л., Генетика підлоги, в збірці: Актуальні питання сучасної генетики, [М.], 1966; Бреславец Л. П., Визначення і спадковість підлоги у вищих рослин, «Тр. по прикладній ботаніці, генетиці і селекції, Сірок. 2», 1934 № 6; Рижков Ст Л., Генетика підлоги, [Хар.], 1936; Лобашев М. Е., Генетика, 2 видавництва, Л., 1967.