Запліднення , сингамія, в рослин, тварин і людини — злиття чоловічою і жіночою статевих кліток — гамет, внаслідок чого утворюється зігота, здатна розвиватися в новий організм. О. лежить в основі статевого розмноження і забезпечує передачу спадкових ознак від батьків нащадкам.
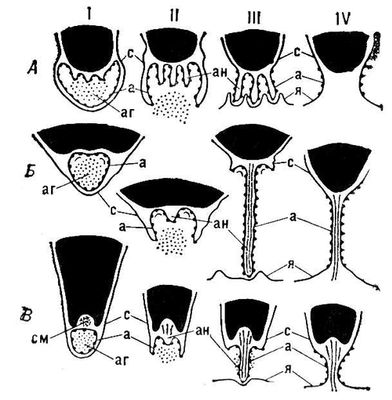
Запліднення в рослин . О. властиве більшості рослин; йому зазвичай передує утворення гаметангиев — статевих органів, в яких розвиваються гамети. Часто ці процеси об'єднують під загальною назвою статевий процес. Рослини, що мають статевий процес, мають в циклі розвитку і мейоз, тобто виявляють зміну ядерних фаз (див. Чергування поколінь ). Типового статевого процесу немає у бактерій і синезелених водоростей; невідомий він і в деяких грибів. Типи статевого процесу в нижчих рослин всілякі. В ряду зелених водоростей він може здійснюватися без утворення гамет, в результаті злиття двох одноклітинних організмів (т.з. гологамія ). Злиття гамет, що мають джгутики, форма і розміри яких однакові, називається ізогамією (см. мал.(малюнок) 1 , 1). Цей тип статевого процесу властивий багатьом водоростям. Одноклітинні водорості (наприклад, деякі хламідомонади) як би самі перетворюються на гаметангиі, утворюючи гамети; в багатоклітинних гаметангиямі стають деякі клітки, що не відрізняються від інших (наприклад, в улотрікса, ульви), або виникають морфологічно відмінні гаметангиі (наприклад, в ектокарпуса). Багато ізогамниє водоростей гетероталлічни: зливаються лише фізіологічно різні (+ і –) гамети (див. Гетероталлізм ). Для водоростей кон'югат (наприклад, спірогіри) характерна кон'югація : протопласт однієї клітки перетікає в іншу (що належить тій же або ін. особини), зливаючись з її протопластом ( мал. 1 , 2). Злиття гамет різної величини (велика — жіноча, менша — чоловіча; наприклад, в деяких хламідомонад), що мають джгутики, називається гетерогамією ( мал. 1 , 3). Злиття крупною безжгутиковою жіночі гамети (яйцеклітина) і дрібною чоловічою, такою, що частіше має джгутики (сперматозоїд), рідше — безжгутиковою (спермаціїв), називається оогамією . Жіночі гаметангиі більшості оогамних нижчих рослин називаються оогоніямі, чоловічі, — антерідіямі. Оогамія характерна для багатьох зелених, діатомових, бурих ( мал. 1 , 4) і всіх червоних водоростей, деяких нижчих грибів. В гологамних, ізо-, гетеро- і багатьох оогамних рослин О. відбувається у воді, в деяких оогамних (вольвокса, вошерії) — у жіночих гаметангиях — оогоніях, до яких сперматозоїди, що вийшли у воду, активно переміщаються (що, мабуть, обумовлене хемотаксисом ), а спермації червоних водоростей — пасивно, струмом води. В рослин з гаметангиогамієй гамети не диференціюються. Так, в мукорових грибів зливаються багатоядерні гаметангиі, що виникають на кінцях виростов міцелієв (різних при гетероталлізме) ( мал. 1 , 5); при цьому попарно зливаються і ядра. Цей тип гаметангиогамії називається зігогамієй . У більшості сумчастих грибів багатоядерний протопласт антерідію переливається в базальну клітку жіночого гаметангия (аскогон), що містить протопласт з безліччю ядер; ядра лише попарно зближуються, утворюючи т.з. дікаріони (перший етап статевого процесу — плазмогамія). З аскогона зростають гіфи, в них ядра дікаріонов синхронно діляться; на кінцях гіф виникають сумки — клітки, що містять по дикаріону. У сумках (аськах) відбувається другий етап статевого процесу — каріогамія, тобто злиття ядер ( мал. 1 , 6). Для базідіальних грибів характерна соматогамія: вони не утворюють ні гамет, ні гаметангиев; плазмогамія відбувається у них при злитті двох одноядерних кліток, т.з. первинних (+ і –) міцелієв; двуядерная клітка, що виникає при цьому, дає початок вторинному міцелію, що складається з кліток, дікаріони, що містять; на цьому міцелії утворюються базидії, в них і відбувається каріогамія ( мал. 1 , 7). Гаметангио- і соматогамія — вироблене грибами в процесі еволюції пристосування до існування поза водним середовищем.
Всі вищі рослини оогамни, але О. у них здійснюється по-різному. Типові гаметангиі вищих рослин — антерідії (чоловічі) і архегонії (жіночі) многоклеточни; клітки зовнішнього шару гаметангия стерильні. Яйцеклітини утворюються в архегоніях по одній, сперматозоїди — в антерідіях, як правило, багато. Мохо- і папоротеподібним для здійснення О. необхідна вода, в якій що вийшли з антерідієв сперматозоїди пливуть до архегоніям. З вершини готового к О, що розкрилася. архегонія виступає слиз, що залучає сперматозоїдів. Рухаючись в слизі сперматозоїди досягають яйцеклітини і один з них зливається з нею ( мал. 1 , 8 і 9). В папоротеподібних і насінних рослин О. відбувається на (або в) заростку (гаметофіте ), що існує в перших самостійно, а в других — на спорофіті . В равноспорових папоротей заростки обоєполи, в різноспорових і всіх насінних рослин раздельнополи. В насінних рослин антерідієв немає: сперматозоїди (в саговників, гинкго) або безжгутикові спермії (у всіх інших) утворюються в чоловічих заростках (пилкових зернах ). В деяких голосеменних (гнетум, вельвічия) і всіх покритосеменних архегонієв немає і яйцеклітини знаходяться в жіночих заростках. В насінних рослин О. можливо лише після запилення — перенесення пилкових зерен з мікроспорангіїв в пилкові камери семезачатков (в голосеменних) або на рильця товкачів (в покритосеменних). В саговників і гинкго сперматозоїди виходять в архегоніальную камеру семезачатка ( мал. 1 , 10) і, рухаючись в рідині, виробленій самою рослиною, досягають архегонієв. В насінних рослин, що мають спермії, останні переміщаються до яйцеклітинам по пилкових трубках ( мал. 1 , 11 і 12). В покритосеменних відбувається подвійне запліднення : один спермій зливається з яйцеклітиною, другою, — з центральною кліткою зародкового мішка (жіночого заростка). Здійснення О. незалежно від наявності вільної води — одне з найважливіших пристосувань насінних рослин до існування на суші.
Літ.: Мейєр До. І., Розмноження рослин, М., 1937; Навашин С. Р. Ізбр. праці, т. 1, М. — Л., 1951; Тахтаджян А. Л., Вищі рослини, т. 1, М. — Л., 1956; Сладков А. Н., Статевий процес і життєві цикли в рослин, «Біологічні науки», 1969 № 3—4.
А. Н. Сладков.
Запліднення в тварин і людини полягає в злитті (сингамії) двох гамет різної підлоги — спермію (сперматозоїда ) і яйця . О. має двояке значення: 1) контакт спермію з яйцем виводить останнє із загальмованого стану і спонукає до розвитку (див. Активація яйця ); 2) злиття гаплоїдіих ядер спермію і яйця — каріогамія — приводить до виникнення диплоїдного синкаріона, об'єднуючого батьківські і материнські спадкові чинники. Виникнення при О. нових комбінацій цих чинників створює генетична різноманітність, що служить матеріалом для природного відбору і еволюції вигляду. Необхідна передумова О. — зменшення числа хромосом удвічі, що відбувається під час мейозу. Ці ділення в чоловічих гамет здійснюються до формування спермію, тоді як співвідношення між діленнями дозрівання яйця і О. у різних тварин різно: спермій може проникати в яйце до початку мейозу (губки, деякі черв'яки, молюски, з ссавців — собака, лисиця, кінь); на стадії метафази 1-го ділення дозрівання (деякі черв'яки, молюски, комахи, асцидії); на стадії метафази 2-го ділення (ланцетник, багато хребетних) і після завершення мейозу (кишечнополостниє, морські їжаки).
Зустріч сперматозоїда з яйцем зазвичай забезпечується плавальними рухами чоловічих гамет після того, як вони виметани у воду або введені в статеві дороги самки (див. Запліднення ). Зустрічі гамет сприяє вироблення яйцями гамонов, що підсилюють рухи сперміїв і що подовжують період їх рухливості, а також речовин, що викликають скупчення сперміїв поблизу яйця. Виникнення таких скупчень в гідроїдних поліпів з роду Campanularia і деяких риб інколи розглядають як наслідок залучення сперміїв (хемотаксис), подібного спостережуваному при О. в мохів, папоротей і ін. Проте існування направлених рухів для сперміїв тварин недоведене; сперматозоїди рухаються безладно і вступають в контакт з яйцем в результаті випадкового зіткнення, а утворення їх скупчень, ймовірно, викликається дією механізму типа «пастки», що затримує спермії що випадково наблизилися до яйця.
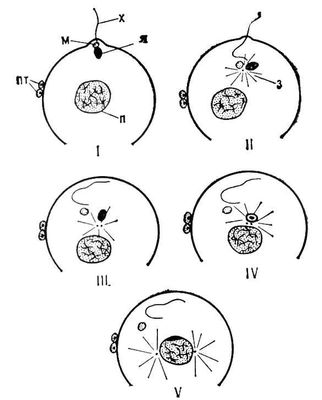
Зріле яйце оточене оболонками, що мають у деяких тварин отвору для проникнення сперміїв, — мікропилі . У більшості тварин мікропилі відсутній, і, щоб досягти поверхні ооплазми, спермії повинні проникнути через оболонку, що здійснюється за допомогою спеціального органоїду сперматазоїда — акросоми . Після того, як спермій кінцем голівки торкнеться яєчної оболонки, відбувається акросомная реакція: акросома розкривається, виділяючи вміст акросомной гранули ( мал. 2 , стадії I, II), і укладені в гранулі ферменти розчиняють яєчні оболонки. У тому місці де розкрилася акросома, її мембрана зливається з плазматичною мембраною спермію; в підстави акросоми акросомная мембрана вигинається і утворює один або декілька виростов ( мал. 2 , стадія II) які заповнюються розташованим між акросомою і ядром (субакросомальним) матеріалом, подовжуються і перетворюються на акросомниє нитки або трубочки ( мал. 2 , стадія III). Довжина цих ниток у різних тварин варіює від 1 до 90 мкм (залежно від товщини бар'єру, який спермію доводиться долати). Акросомная нитка проходіт через розчинену зону яєчної оболонки, вступає в контакт з плазматичною мембраною яйця і зливається з нею ( рис 2 , стадії III, IV). У тварин, спермії яких проникають в яйце через мікропилі (комахи, головоногі молюски, осетрові і костисті риби), акросома втрачає своє первинне значення і інколи редукується або повністю зникає (в деяких веснянок, костистих риб). У ссавців овуліровавшєє яйце, окрім оболонки, оточено декількома шарами фолікулярних кліток яйцеродного горбка. У коня, корови, вівці фолікулярні клітки розсіваються незабаром після овуляції, і спермій вільно досягає поверхні яєчної оболонки. У більшості ссавців клітки яйцеродного горбка зберігаються впродовж декількох годинників і, щоб проникнути через цей бар'єр, спермії виділяють фермент гиалуронідазу, який розчиняє речовину, що зв'язує фолікулярні клітки між собою. Гіалуронидаза, як і фермент, розчинювальний яєчну оболонку, поміщена в акросомі. Відразу після еякуляції спермії нездібні до виділення цих ферментів; така здатність виникає під дією вмісту жіночих статевих доріг, що викликає певні фізіологічні зміни сперміїв (процес капацитациі).
З моменту злиття плазматичних мембран гамет в місці контакту акросомной нитки з поверхнею ооплазми яйце і спермій — вже єдина клітка — зігота. Незабаром виявляються перші ознаки активації яйця: кортікальная реакція і стягання ооплазми в місці контакту з акросомной ниткою спермію, що приводить до утворення сприймаючого горбка. Ооплазма цього горбка обтікає ядро, центріолі і мітохондрії сперматозоїда, а інколи і осьовий стрижень його хвоста, залучаючи їх в глиб яйця, тоді як плазматична мембрана спермію залишається на поверхні і вбудовується в плазматичну мембрану яйця, так що поверхнева мембрана зіготи має мозаїчну будову. Занурившись в ооплазму, голівка спермію повертається на 180°, і в її підстави формується сперматічеськая зірка ( мал. 3 , стадії I, II). Поступово голівка набухає і перетвориться в бульбашковидний чоловічий пронуклеус, що переміщається услід за сперматічеськой зіркою, яка як би захоплює його за собою ( мал. 3 , стадії II, III, IV). Чоловічий пронуклеус зближується з жіночим, а сперматічеськая зірка ділиться на дві, що беруть участь потім в утворенні веретена 1-го ділення дроблення. В кишечнополостних, плоских черв'яків, морських їжаків пронуклеуси зливаються в єдине ядро зіготи ( мал. 3 , стадія V), у деяких круглих черв'яків, молюсків, ракоподібних, риб і земноводних вони тривалий час залишаються в тісному контакті, але не зливаються, і об'єднання батьківського і материнського ядерного матеріалу відбувається лише на стадії метафази 1-го ділення дроблення. Одночасно з цими змінами в яйці підвищується інтенсивність обміну речовин: збільшується проникність клітинної мембрани, активується синтез білка і ін.
При О. яєць тварин із зовнішнім заплідненням в яйце проникає лише один спермій (фізіологічна моноспермії); це забезпечується особливим механізмом, в основі якого лежить процес секреції вмісту кортікальних телець, що запобігає проникненню сперміїв в ооплазму. Серед тварин з внутрішнім заплідненням поряд з моноспермнимі зустрічаються і такі, в яких в яйце проникає декілька сперміїв (фізіологічна поліспермія ); проте і в цих випадках з жіночим пронуклеусом зливається лише одне сперматічеськоє ядро. Див. також Спадковість і Пів .
Літ.: Ротшильд Н. М., Запліднення, пер.(переведення) з англ.(англійський), М., 1958; Дорфман Ст А., Физико-хімічні основи запліднення, М., 1963; Гинзбург А. С., Запліднення у риб і проблема поліспермії, М., 1968; Austin С. R., Fertilization, Englewood Cliffs (N. J.), 1965; Fertilization, ed. Ch. B. Metz, A. Monroy, v. 1—2, N. Y. — L., 1967—69; Reproduction in mammals, ed. C. R. Austin, R. V. Short, book 1 — Germ cells and fertilization, L., 1972.