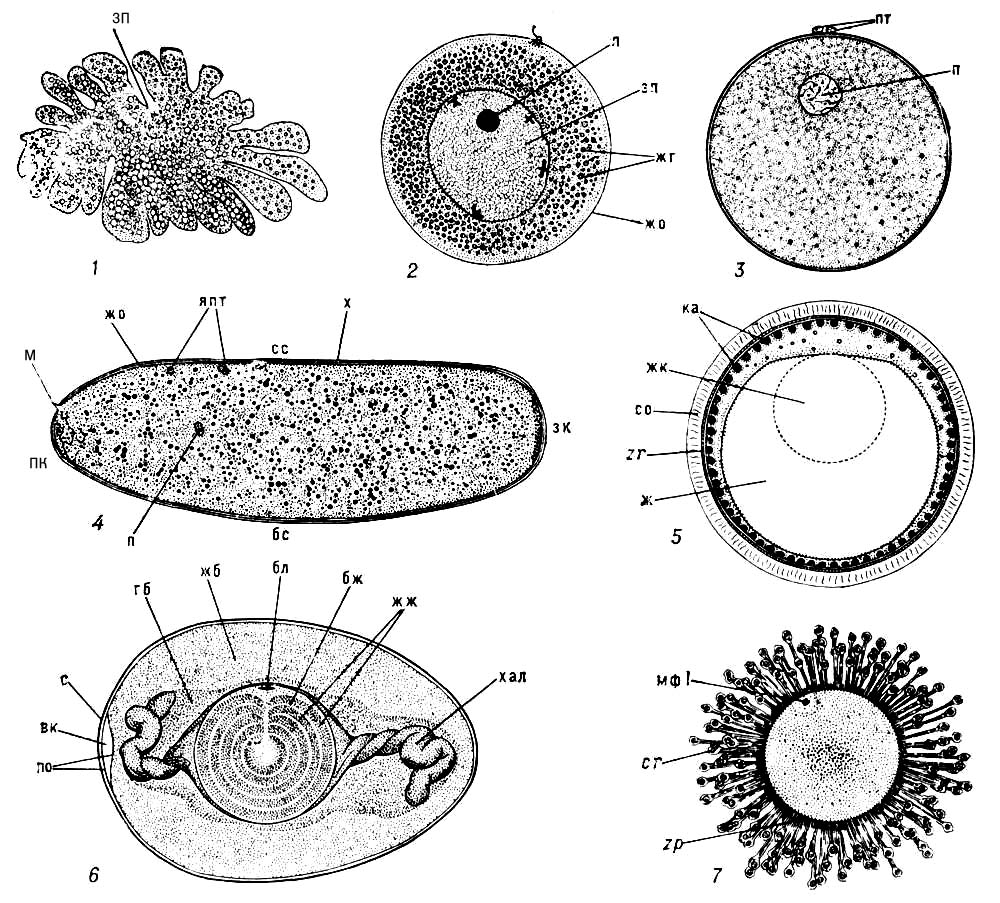
Яйце , жіноча статева клітка людини, тварин і рослин (яйцеклітина), з якої розвивається новий організм в результаті запліднення або дорогою партеногенезу . Я. людини і тварин — високоспеціалізована клітка, що містить запаси живильних речовин, необхідних для розвитку зародка. Я. буває, як правило, одягнене яєчними оболонками . Формування Я. (оогенез ) зазвичай відбувається в яєчниках. У більшості тварин Я. має округлу або овальну форму ( мал. 2 , 3 , 5—7 ), рідше, наприклад у комах ( мал. 4 ), головоногих молюсків, — подовжену. В найбільш примітивних багатоклітинних тварин (губки, деякі кишечнополостниє) Я. не має певної форми ( мал. 1 ) і здібно до амебоїдних рухів. Розміри Я. варіюють залежно від кількості жовтка в цитоплазмі. Так, бідні жовтком Я. ссавців плацентарних мають діаметр (без оболонок) від 50 мкм (полівка) або 70 мкм (щур, миша) до 140—150 мкм (кажан, морська свинка, корова, собака), максимально — 180 мкм (вівця); діаметр Я. людини — 89—91 мкм . Схожі розміри мають Я. багатьох безхребетних — кишечнополостних, черв'яків, голкошкірих, молюсків. При накопиченні великих запасів жовтка Я. можуть досягати крупних, інколи гігантських розмірів: Я. деякі безхребетні мають діаметр близько 1,4 мм (деякі молюски, голкошкірі, ракоподібні), качкодзьоба — 4,3 мм , лососевих риб — 7—9 мм , акулообразних — 50—70 мм ; дуже великі також Я. деяких птиць: діаметр власне Я. курки (без тієї, що оточує його білкової оболонки) понад 30 мм , Я. страуса — 80 мм (при загальній довжині Я. у шкаралупі 155 мм ). Між величиною Я. і розмірами тіла дорослого організму немає чіткої залежності; Я. людину і кролика мають однакові розміри, у акваріумної рибки гуппі, при довжині самки 5—6 мм , Я. удвічі більше, ніж у півтораметрового блакитного тунця (діаметр Я. відповідно 2—2,1 і 1—1,1 мм ). Зазвичай спостерігається зворотна кореляція між числом Я. і їх розмірами. Зокрема, тварини, що охороняють своє потомство, виробляють, як правило, трохи великих Я., наприклад птиці відкладають від 1 (пінгвіни, деякі хижаки) до 22 (куріпка) Я. великого розміру, морські сомики з сімейства Ariidae, що виношують Я. у ротовій порожнині, виробляють по 20—40 Я. діаметром 17—21 мм (найбільші Я., відомі у костистих риб). В той же час у риб що викидають ікру у воду і не проявляють турботи про потомство, плодючість дуже велика: багато коропових, наприклад вобла, лящ, карась, викидають до 300 тис., сазан — до 1,5 млн. і тріск — до 10 млн. Я. діаметром менше 2 мм.
У деяких тварин спостерігається поліморфізм Я. Зокрема, у тварин із складним циклом розмноження — коловерток, ветвістоусих рачків, тлею — спостерігається сезонний поліморфізм Я., наприклад в дафній влітку змінявся декілька поколінь самок, що виробляють «літні» Я. з малою кількістю жовтка і тонкою оболонкою, які розвиваються без запліднення; при настанні несприятливих умов самки відкладають багаті жовтком запліднені «зимові» Я., увязнені в захисну оболонку, які продовжують розвиватися лише після тривалого періоду спокою. В коловерток відомо 3 типи Я.: диплоїдні, такі, що дають партеногенетичним шляхом самок; дрібні гаплоїдниє, розвиваються в самців; що покояться, або «зимові», Я., що розвиваються в самок після запліднення. Існує також статевий диморфізм Я.: в метеликів, ручейников, плазунів, птиць. Я., що розвиваються в самця і самку, розрізняються по статевих хромосомах (жіноча гетерогаметія); при зовні помітному статевому диморфізмі (дімегалії) в коловерток, деяких черв'яків великі Я. дають самок, а дрібні — самців.
Зріле Я. володіє певною організацією, яка виявляється в першу чергу в його полярності: часто ядро розташовується поблизу від бідного жовтком (анімального) полюса, тоді як жовткові включення концентруються ближче до протилежного (вегетативному) полюса. Крім того, в Я. багатьох тварин виявляються локальні відмінності (див. Сегрегація ооплазматічеськая), яскраво виражені у молюсків, черв'яків і асцидій. У цих тварин доля різних областей Я. вже визначена і при його розділенні на частини з них розвиваються потворні зародки, в яких відсутні ті або інші органи (мозаїчні Я.). У інших тварин, зокрема в голкошкірих, організація Я. більш лабільна: при його розділенні на частини можуть розвинутися декілька маленьких, але повноцінних зародків (регуляції Я.). Раніше мозаїчні і регуляції Я. протиставляли один одному, проте відмінності між ними непринципові і обумовлені більш менш раннім диференціюванням цитоплазми.
Поверхня Я. одягнена напівпроникною еластичною плазматичною мембраною, скоротністю, що володіє. Поверхневий (кортікальний) шар цитоплазми утворює багаточисельні вирости — мікроворсинки. У цьому шарі у багатьох тварин поміщені особливі тільця, вміст яких багато мукополісахарідамі; такі тільця діаметром від десятих доль мкм (кортікальниє гранули двостулкових молюсків, голкошкірих, ссавців) до 5—40 мкм (кортікальниє альвеоли костистих риб, мал.(малюнок) 5) розташовуються в один або декілька рядів поблизу плазматичної мембрани. При активації яйця їх вміст виділяється з цитоплазми під оболонку (див. Кортікальная реакція ) . Недалеко від поверхні зазвичай лежать і пігментні гранули. Постійними компонентамі цитоплазми Я., так само як і інших кліток, є мітохондрії, комплекси Гольджі (які у багатьох тварин концентруються на периферії Я.), ендоплазматична мережа і рибосоми. Клітинний центр в Я. більшості тварин до кінця оогенезу зникає і лише зрідка (наприклад, у деяких п'явок, планарій і ін.) зберігається, виявляючись під час ділень дозрівання. У останніх випадках після запліднення в Я. формується новий клітинний центр — за рахунок центріолей, що вносяться сперматозоїдом або, рідко, шляхом новоутворення. Специфічним компонентом цитоплазми Я. є жовткові включення; відмінності в кількості жовтка і його розподілі у різних тварин обумовлюють різних типів дроблення Я. У цитоплазмі міститься великий запас макромолекул (вся синтезуюча для білка система, включаючи інформаційну РНК(рибонуклеїнова кислота) і необхідні ферменти, а також ДНК(дезоксирибонуклеїнова кислота)-и РНК(рибонуклеїнова кислота) -полімерази, негистоновиє ядерні білки — тубулін і ін.), які використовуються лише після активації яйця і на ранніх стадіях розвитку зародка. ПРО Я. рослин див.(дивися) в ст. Яйцеклітина .
Літ.: Вільсон Е., Клітка і її роль в розвитку і спадковості, пер.(переведення) з англ.(англійський), т. 1—2, М. — Л., 1936—40; Соколів І. І., Цитологичеськие основи статевого розмноження багатоклітинних тварин, в кн.: Керівництво по цитології, т. 2, М. — Л., 1966, с. 390—460; Бодемер Ч., Сучасна ембріологія, пер.(переведення) з англ.(англійський), М., 1971; Boyd J. D., Hamilton W. J., Cleavage, early development and implantation of the egg, в кн.: Marshall''s physiology of reproduction, v. 2, L., 1952; Austin C. R., The mammalian egg, Oxf., 1961.