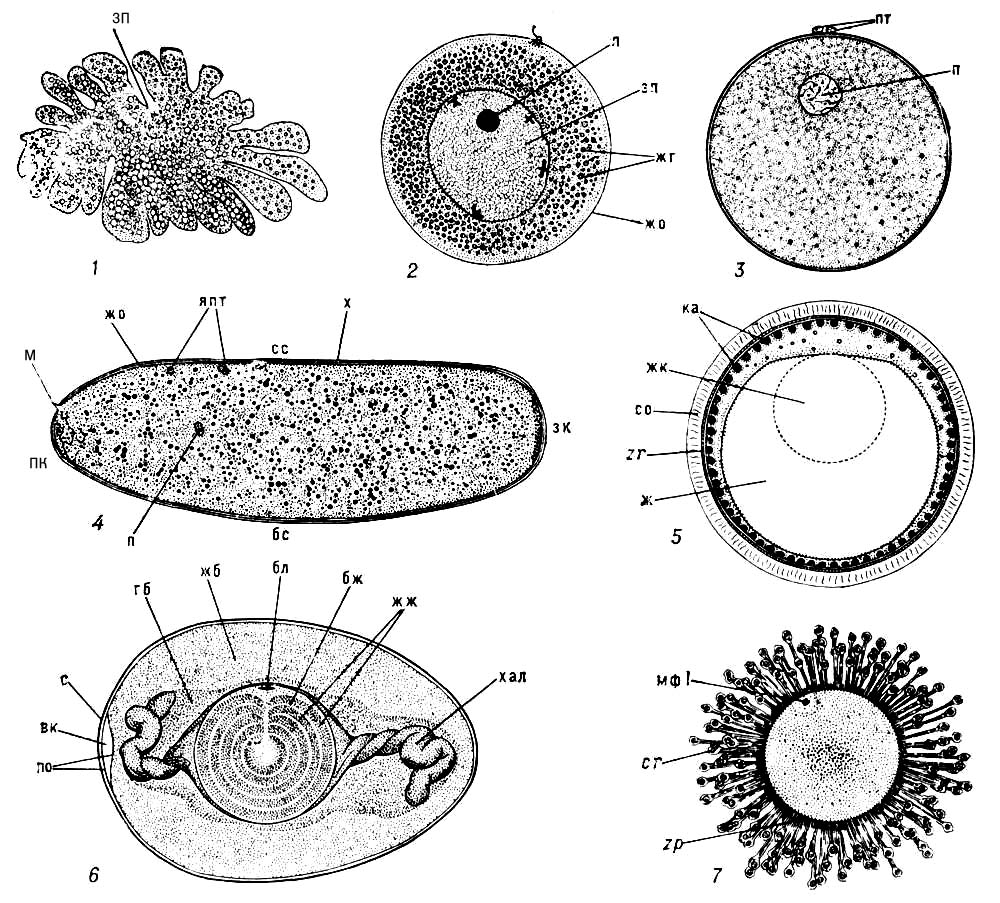
Яйцо, женская половая клетка человека, животных и растений (яйцеклетка), из которой развивается новый организм в результате оплодотворения или путём партеногенеза. Я. человека и животных — высокоспециализированная клетка, содержащая запасы питательных веществ, необходимых для развития зародыша. Я. бывает, как правило, одето яйцевыми оболочками. Формирование Я. (оогенез) обычно происходит в яичниках. У большинства животных Я. имеет округлую или овальную форму (рис. 2, 3, 5—7), реже, например у насекомых (рис. 4), головоногих моллюсков, — удлинённую. У наиболее примитивных многоклеточных животных (губки, некоторые кишечнополостные) Я. не имеет определенной формы (рис. 1) и способно к амёбоидным движениям. Размеры Я. варьируют в зависимости от количества желтка в цитоплазме. Так, бедные желтком Я. плацентарных млекопитающих имеют диаметр (без оболочек) от 50 мкм (полёвка) или 70 мкм (крыса, мышь) до 140—150 мкм (летучая мышь, морская свинка, корова, собака), максимально — 180 мкм (овца); диаметр Я. человека — 89—91 мкм. Сходные размеры имеют Я. многих беспозвоночных — кишечнополостных, червей, иглокожих, моллюсков. При накоплении больших запасов желтка Я. могут достигать крупных, иногда гигантских размеров: Я. некоторых беспозвоночных имеют диаметр около 1,4 мм (некоторые моллюски, иглокожие, ракообразные), утконоса — 4,3 мм, лососёвых рыб — 7—9 мм, акулообразных — 50—70 мм; очень велики также Я. некоторых птиц: диаметр собственно Я. курицы (без окружающей его белковой оболочки) свыше 30 мм, Я. страуса — 80 мм (при общей длине Я. в скорлупе 155 мм). Между величиной Я. и размерами тела взрослого организма нет чёткой зависимости; Я. человека и кролика имеют одинаковые размеры, у аквариумной рыбки гуппи, при длине самки 5—6 мм, Я. вдвое крупнее, чем у полутораметрового голубого тунца (диаметр Я. соответственно 2—2,1 и 1—1,1 мм). Обычно наблюдается обратная корреляция между числом Я. и их размерами. В частности, животные, охраняющие своё потомство, производят, как правило, немного крупных Я., например птицы откладывают от 1 (пингвины, некоторые хищники) до 22 (куропатка) Я. большого размера, морские сомики из семейства Ariidae, вынашивающие Я. в ротовой полости, производят по 20—40 Я. диаметром 17—21 мм (самые крупные Я., известные у костистых рыб). В то же время у рыб, вымётывающих икру в воду и не проявляющих заботы о потомстве, плодовитость очень велика: многие карповые, например вобла, лещ, карась, вымётывают до 300 тыс., сазан — до 1,5 млн. и треска — до 10 млн. Я. диаметром менее 2 мм.
У некоторых животных наблюдается полиморфизм Я. В частности, у животных со сложным циклом размножения — коловраток, ветвистоусых рачков, тлей — наблюдается сезонный полиморфизм Я., например у дафний летом сменяется несколько поколений самок, производящих «летние» Я. с малым количеством желтка и тонкой оболочкой, которые развиваются без оплодотворения; при наступлении неблагоприятных условий самки откладывают богатые желтком оплодотворённые «зимние» Я., заключённые в защитную оболочку, которые продолжают развиваться только после длительного периода покоя. У коловраток известны 3 типа Я.: диплоидные, дающие партеногенетическим путём самок; мелкие гаплоидные, развивающиеся в самцов; покоящиеся, или «зимние», Я., развивающиеся в самок после оплодотворения. Существует также половой диморфизм Я.: у бабочек, ручейников, пресмыкающихся, птиц. Я., развивающиеся в самца и самку, различаются по половым хромосомам (женская гетерогаметия); при внешне различимом половом диморфизме (димегалии) у коловраток, некоторых червей крупные Я. дают самок, а мелкие — самцов.
Зрелое Я. обладает определенной организацией, которая проявляется в первую очередь в его полярности: часто ядро располагается вблизи от бедного желтком (анимального) полюса, тогда как желточные включения концентрируются ближе к противоположному (вегетативному) полюсу. Кроме того, в Я. многих животных обнаруживаются локальные различия (см. Сегрегация ооплазматическая), ярко выраженные у моллюсков, червей и асцидий. У этих животных судьба разных областей Я. уже определена и при его разделении на части из них развиваются уродливые зародыши, у которых отсутствуют те или иные органы (мозаичные Я.). У других животных, в частности у иглокожих, организация Я. более лабильна: при его разделении на части могут развиться несколько маленьких, но полноценных зародышей (регуляционные Я.). Раньше мозаичные и регуляционные Я. противопоставляли друг другу, однако различия между ними непринципиальные и обусловлены более или менее ранней дифференцировкой цитоплазмы.
Поверхность Я. одета полупроницаемой эластичной плазматической мембраной, обладающей сократимостью. Поверхностный (кортикальный) слой цитоплазмы образует многочисленные выросты — микроворсинки. В этом слое у многих животных заключены особые тельца, содержимое которых богато мукополисахаридами; такие тельца диаметром от десятых долей мкм (кортикальные гранулы двустворчатых моллюсков, иглокожих, млекопитающих) до 5—40 мкм (кортикальные альвеолы костистых рыб, рис.(рисунок) 5) располагаются в один или несколько рядов вблизи плазматической мембраны. При активации яйца их содержимое выделяется из цитоплазмы под оболочку (см. Кортикальная реакция). Недалеко от поверхности обычно лежат и пигментные гранулы. Постоянными компонентами цитоплазмы Я., так же как и других клеток, являются митохондрии, комплексы Гольджи (которые у многих животных концентрируются на периферии Я.), эндоплазматическая сеть и рибосомы. Клеточный центр в Я. большинства животных к концу оогенеза исчезает и лишь изредка (например, у некоторых пиявок, планарий и др.) сохраняется, обнаруживаясь во время делений созревания. В остальных случаях после оплодотворения в Я. формируется новый клеточный центр — за счёт центриолей, вносимых сперматозоидом или, редко, путём новообразования. Специфическим компонентом цитоплазмы Я. являются желточные включения; различия в количестве желтка и его распределении у разных животных обусловливают разные типы дробления Я. В цитоплазме содержится большой запас макромолекул (вся белок-синтезирующая система, включая информационную РНК(рибонуклеиновая кислота) и необходимые ферменты, а также ДНК(дезоксирибонуклеиновая кислота)-и РНК(рибонуклеиновая кислота)-полимеразы, негистоновые ядерные белки — тубулин и др.), которые используются только после активации яйца и на ранних стадиях развития зародыша. О Я. растений см.(смотри) в ст. Яйцеклетка.
Лит.: Вильсон Э., Клетка и ее роль в развитии и наследственности, пер.(перевод) с англ.(английский), т. 1—2, М. — Л., 1936—40; Соколов И. И., Цитологические основы полового размножения многоклеточных животных, в кн.: Руководство по цитологии, т. 2, М. — Л., 1966, с. 390—460; Бодемер Ч., Современная эмбриология, пер.(перевод) с англ.(английский), М., 1971; Boyd J. D., Hamilton W. J., Cleavage, early development and implantation of the egg, в кн.: Marshall's physiology of reproduction, v. 2, L., 1952; Austin C. R., The mammalian egg, Oxf., 1961.