Дроблення яйця, його сегментація, ряд послідовних ділень яйця, в результаті яких воно розділяється на все дрібніші клітки (бластомери ). Д. — неодмінна стадія розвитку всіх багатоклітинних тварин. Зазвичай починається після зближення чоловічого і жіночого пронуклеусов (див. Запліднення ) і об'єднання їх хромосом на веретені 1-го ділення Д. У деяких тварин відбувається Д. незапліднених яєць (див. Партеногенез ). Інколи запліднені яйця знаходяться деякий час у спокої (див. Діапауза ) і спонукають до розвитку зміною зовнішніх умов (наприклад, температури довкілля). Спочатку в період синхронних ділень, ядра у всіх бластомерах діляться з однаковим і постійним ритмом, ядерний цикл короткий; в різних груп тварин цей період неоднаковий за тривалістю, а у ссавців відсутній. Потім, в період асинхронних ділень, або бластуляциі, ядерний цикл подовжується, порушується синхронність в діленні різних ядер, на стадії інтерфази в них починається синтез рибонуклеїнової кислоти (РНК), виявляється їх морфогенетична функція. Розділення цитоплазми (цитотомія) слідує за діленням ядер (каріотомія), але, як правило, відстає від нього. Д. не супроводиться зростанням, і зародок зберігає вихідні розміри яйця. Після закінчення Д. зародок досягає стадії бластули .
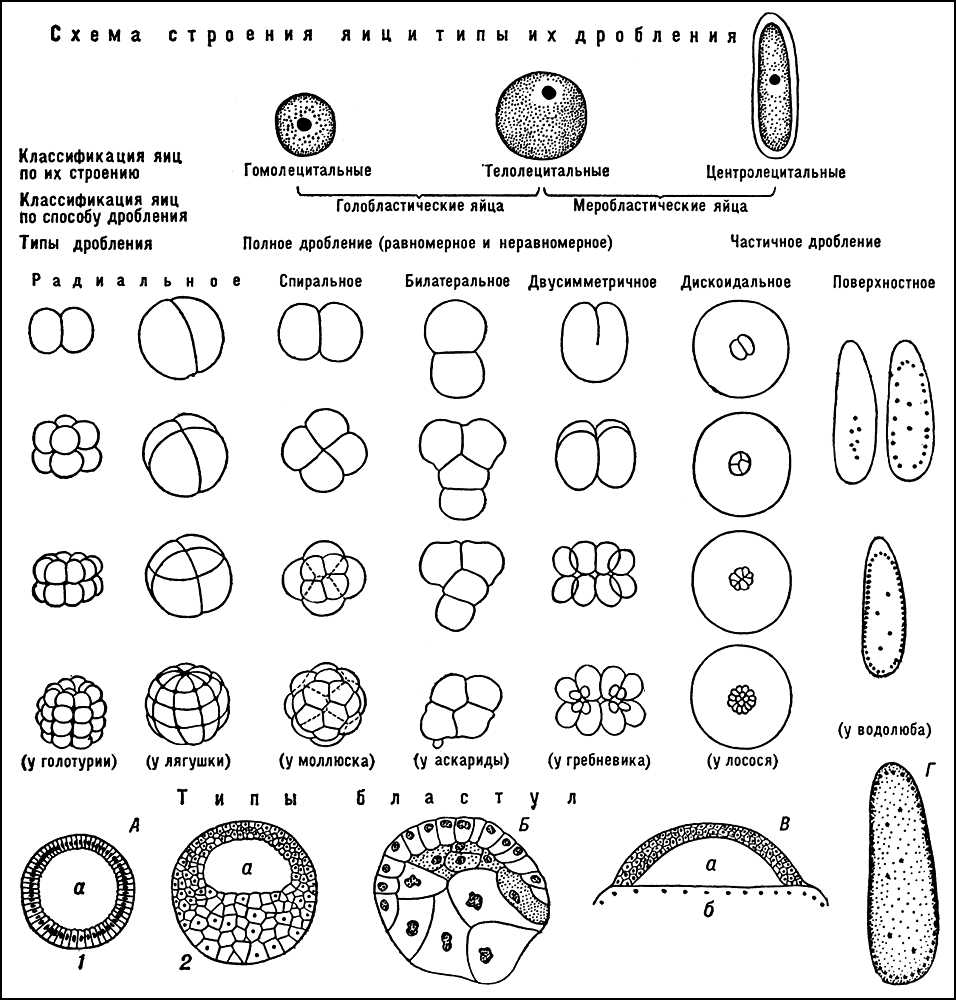
На характер Д. впливають кількість і розподіл жовтка в цитоплазмі яєць. Що містять порівняно мало рівномірно розподіленого жовтка гомолецитальниє яйця зазнають повне рівномірне Д. Чаще жовток розподілений в цитоплазмі яйця нерівномірно (телоцитальні і центролецитальниє яйця). Область, що містить більше жовтка, ділиться повільніше бідною жовтком — повне нерівномірне Д., або зовсім не ділиться — часткове Д. Яйця, що зазнають повне Д., називають голобластічеськимі, часткове Д., — меробластічеськимі. До голобластічеським відносяться гомолецитальниє (наприклад, яйця багатьох безхребетних, ланцетника, ссавців) і частина телоцитальних (наприклад, яйця деяких членистоногих, більшості земноводних), які зазнають повне, але нерівномірне Д. (дрібні бластомери називаються мікрозаходами, середні — мезомерами, великі — макрозаходами). До меробластічеським яєць відносяться частина телоцитальних і центролецитальниє з великою кількістю жовтка. У таких телоцитальних яйцях ділиться лише бідна жовтком анімальная частина яйця, яка послідовно розділяється на 2, 4 і більше число бластомерів, створюючих диск кліток на поверхні жовтка, що не дробиться, — дискоїдальне Д. Оно характерний для яєць скорпіонів, головоногих молюсків, акулових і костистих риб, птиць, плазуючих і нижчих ссавців. В результаті дискоїдального Д. утворюється діськобластула, порожнина якої обмежена розмірами бластодерми. Часткове Д. характерний і для центролецитальних яєць більшості членистоногих. Після запліднення ядро починає ділитися. Після декількох синхронних ділень ядра з цитоплазмою, що оточує їх, переміщаються по цитоплазматичних містках в поверхневий шар цитоплазми, який спочатку є симпласт, потім довкола кожного ядра відособляється окрема клітка. В результаті утворюється зародок, стінка якого складається з одного шару кліток (бластодерми), а центральна частина зайнята жовтком, що не розділився, з клітками, що знаходяться в нім (вітеллофагамі); такий зародок називається перибластулою, а Д. — поверхневим, або синцитіальним.
На характер Д. впливають і властивості цитоплазми яйця, що визначають положення веретен ділення і, як наслідок, положення бластомерів один відносно одного, оскільки плоскість дроблення завжди перпендикулярна осі веретена. За ознакою відносного положення бластомерів при повному Д. розрізняють радіальне, спіральне, білатеральне і двусимметрічноє Д. Прі радіальному Д., властивому багатьом кишково-порожнинним, голкошкірим, земноводним і ін., бластомери розташовуються так, що будь-яка плоскість, яку можна провести через анімально-вегетатівну вісь яйця, є плоскістю симетрії. Перші 2 борозни проходят зазвичай меридіональний, а 3-я — екваторіально; потім відбувається чергування меридіональних і екваторіальних ділень. В результаті радіального Д. утворюється багатоклітинна бульбашка з порожниною — целобластула.
При спіральному Д., характерному для більшості турбеллярій, кольчецов, немертін, молюсків і ін., мікрозаходи, що відділяються від перших 4 бластомерів (макромеров), розташовуються в проміжках між ними. Відбуваються зсуви бластомерів верхнього ярусу відносно ніжнего управо — дексиотропноє Д., або вліво — леотропноє Д. Прі спіральному Д. зародок на стадії бластули має порожнину (нерівномірна целобластула) або не має її (стерробластула). При білатеральному Д. (у круглих черв'яків асцидій), а також на пізніх стадіях спірального Д. ділення відбуваються так, що у зародків є лише одна плоскість симетрії. Двусимметрічноє Д. спостерігається дуже рідко (гребневіки) і характеризується наявністю двох плоскості симетрії. Див. схему будови яєць, типів їх дроблення і типів бластул . Той або інший тип Д. зазвичай властивий більшості представників того або іншого класу тварин, але інколи в межах класу спостерігаються різні типи Д. Так серед земноводних, більшості яких властиве повне нерівномірне Д., в безногих земноводних є дискоїдальне Д.; у ссавців відбувається як дискоїдальне (однопрохідні), так і повне Д. (всі вищі ссавці). Останнє по ряду ознак (відособлення зародкового диска і позазародкової частини) наближається до дискоїдального, від якого воно сталося. В результаті повного Д. виникає бластоциста; частина її стінки, представлена щільним скупченням кліток утворює зародковий диск, остання частина є трофобласт .
В процесі Д. ядра діляться рівномірно (ядра всіх бластомерів несуть повний об'єм генетичної інформації і рівноцінні як один одному, так і ядру зіготи), а цитоплазма ділиться нерівномірно. Відмінності у властивостях цитоплазми перших бластомерів у різних тварин виражені в неоднаковій мірі і залежать від рівня диференціювання її в оогенезі (див. Сегрегація ооплазматічеськая). У одних тварин при штучному розділенні двох перших бластомерів з кожного утворюється цілий зародок, в інших — лише його частина, т.к. в яйцях різних тварин на початок Д. цитоплазма досягає різної міри диференціювання (найбільш раннє диференціювання характерна для яєць із спіральним, білатеральним і поверхневим Д.). На цій підставі інколи розрізняють яйця регуляцій і мозаїчних.
В процесі Д. рівноцінні по генотипу ядра приходять у взаємодію з цитоплазмою, що якісно розрізняється в різних бластомерах, що є умовою диференціальної реалізації в них генетичної інформації (див. Зародковий розвиток ).
Літ.: Іванов П. П., Керівництво по загальній і порівняльній ембріології, Л., 1945; Токин Би. П., Загальна ембріологія, [2 видавництва], М., 1970.