Чергування поколінь, закономірна зміна в організмів генерацій, що розрізняються типом розмноження.
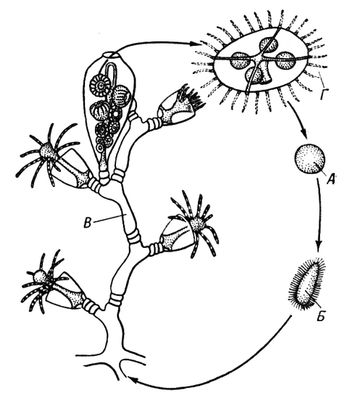
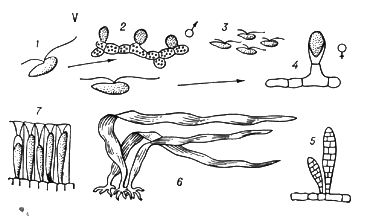
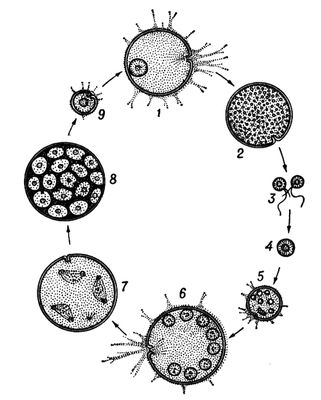
У тварин є первинне і вторинне Ч. п. Первинним Ч. п., властивим багатьом простим, вважають зміну статевої генерації поколінням, що розмножується нестатевими клітками (агаметамі). Так, у форамініфер ( мал. 1 ) покоління, що чергуються, представлені статевими і безстатевими особинами — гамонтамі і агамонтамі. Гамонти в результаті багатократного ділення ядра утворюють гамети, які, копуліруя попарно, дають зіготу, що зростає в агамонта. Останній в результаті шизогонії розпадається на агамети — майбутні гамонти. Редукційне ділення (мейоз ) відбувається перед утворенням агамет, тому статеве покоління гаплоїдно, так само як і гамети, тоді як зігота і агамонти діплоїдни. В споровіков і жгутіконосцев діплоїдна лише зігота, оскільки мейоз здійснюється при її першому діленні. В соняшників, деяких жгутіконосцев і інфузорій мейоз пов'язаний з утворенням гамет, які є єдиною гаплоїдной стадією життєвого циклу. Такі ж стосунки властиві всім багатоклітинним тваринам. Вторинне Ч. п. зустрічається у тварин в двох формах. Чергування різних форм статевого розмноження наприклад нормального статевого процесу з партеногенезом, називається гетерогонієй, а чергування статевого розмноження з безстатевим за допомогою багатоклітинних вегетативних тіл або шляхом поперечного ділення — метагенезом. Гетерогонія характерна для трематод, деяких круглих черв'яків і коловерток, а також для ряду членистоногих — дафній, тлею, горіхотворок, деяких мух-галиць і ін. Метагенез дуже характерний для покривників (сальп, боченочников, асцидій, піросом) і кишечнополостних (гідроїдних і сцифоїдних), в яких статеве покоління представлене одіночнимі свободноплавающимі медузами, а безстатеве — сидячими поліпами, незрідка створюючими колонії ( мал. 2 ). До метагенезу в широкому сенсі слід зараховувати і поліембріонію, т.к. размножающиеся вегетативно зародки по суті є недорозвиненим безстатевим поколінням.
Літ.: Мясоєдов С. Ст, Явища розмноження і підлоги на органічному світі, Томськ, 1935; Гартман М., Загальна біологія, пер.(переведення) з йому.(німецький), 2 видавництва, М.— Л., 1936; Догель Ст А., Зоологія безхребетних, 6 видавництво, М., 1975.
А. Ст Іванов.
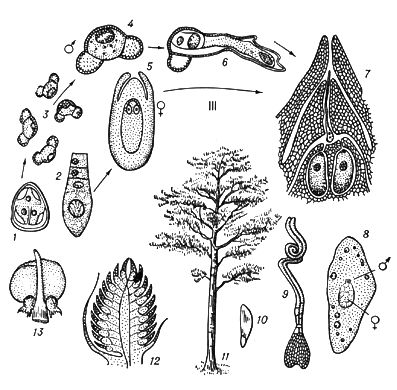
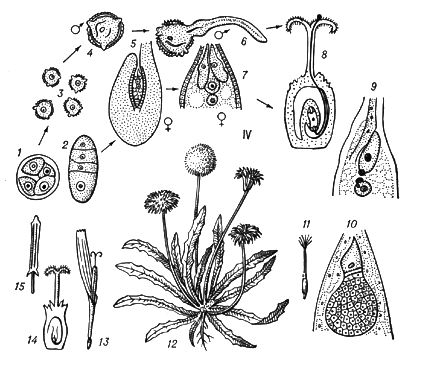
В рослин під Ч. п. зазвичай розуміють чергування в циклах розвитку диплоїдної і гаплоїдной фаз ( мал. 3 ).
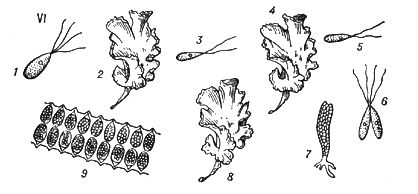
Типове Ч. п. характерний для рослин, в яких многоклеточни як диплоїдна фаза (діплонт), так і гаплоїдная (гаплонт). Діплонт утворює спорангії, в яких в результаті мейозу виникають спори (тому діплонт наз.(назив) також спорофітом ), а гаплонт — гаметангиі, в яких без редукційного ділення утворюються гамети (гаплонт наз.(назив) також гаметофітом ); спорофіт розвивається із зіготи, а гаметофіт — із спори. В одних рослин (наприклад, у водоростей ульви, діктіоти) спорофіт і гаметофіт розвинені однаково, а в ін. домінує або гаметофіт (деякі бурі водорості, наприклад кутлерія, всі мохоподібні), або спорофіт (деякі бурі водорості, наприклад ламінарія, всі папоротеподібні і насінні рослини). В багатьох зелених і, можливо, деяких червоних водоростей діплоїдни лише зіготи, мейотічеськи, що діляться, а в сифонових, діатомових і деяких бурих водоростей (як і у величезної більшості тварин) гаплоїдни лише гамети. В цих рослин фактично Ч. п. немає, хоча зміна ядерних фаз відбувається.
Спорофіти мохоподібних, т.з. спорогонії, розвиваються на гаметофітах. Гаметофіти папоротеподібних (заростки) існують самостійно, а насінних рослин — розвиваються на спорофітах. Заростки ізоспорових (равноспорових) рослин обоєполи, гетероспорових — раздельнополи і більш зредуковані (особливо чоловічі), ніж заростки ізоспорових. Так, в покритосеменних чоловічий заросток — пилкове зерно, жіночий — зародковий мішок. Див. також Запліднення в рослин.
Літ.: Тахтаджян А. Л., Вищі рослини, т. 1 — От псилофітових до хвойних, М-код.—Л., 1956; Поддубная-Арнольді Ст А., Цитоембріология покритосеменних рослин, М., 1976.