Чередование поколений, закономерная смена у организмов генераций, различающихся типом размножения.
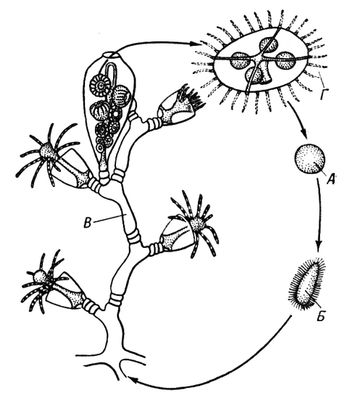
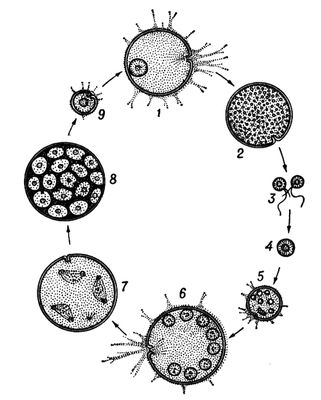
У животных имеются первичное и вторичное Ч. п. Первичным Ч. п., свойственным многим простейшим, считают смену половой генерации поколением, размножающимся неполовыми клетками (агаметами). Так, у фораминифер (рис. 1) чередующиеся поколения представлены половыми и бесполыми особями — гамонтами и агамонтами. Гамонты в результате многократного деления ядра образуют гаметы, которые, копулируя попарно, дают зиготу, вырастающую в агамонта. Последний в результате шизогонии распадается на агаметы — будущие гамонты. Редукционное деление (мейоз) происходит перед образованием агамет, поэтому половое поколение гаплоидно, так же как и гаметы, тогда как зигота и агамонты диплоидны. У споровиков и жгутиконосцев диплоидна только зигота, поскольку мейоз осуществляется при её первом делении. У солнечников, некоторых жгутиконосцев и инфузорий мейоз связан с образованием гамет, которые являются единственной гаплоидной стадией жизненного цикла. Такие же отношения свойственны всем многоклеточным животным. Вторичное Ч. п. встречается у животных в двух формах. Чередование различных форм полового размножения, например нормального полового процесса с партеногенезом,называется гетерогонией, а чередование полового размножения с бесполым посредством многоклеточных вегетативных тел или путём поперечного деления — метагенезом. Гетерогония характерна для трематод, некоторых круглых червей и коловраток, а также для ряда членистоногих — дафний, тлей, орехотворок, некоторых мух-галлиц и др. Метагенез очень характерен для оболочников (сальп, бочёночников, асцидий, пиросом) и кишечнополостных (гидроидных и сцифоидных), у которых половое поколение представлено одиночными свободноплавающими медузами, а бесполое — сидячими полипами, нередко образующими колонии (рис. 2). К метагенезу в широком смысле следует причислять и полиэмбрионию, т.к. размножающиеся вегетативно зародыши по существу представляют собой недоразвитое бесполое поколение.
Лит.: Мясоедов С. В., Явления размножения и пола в органическом мире, Томск, 1935; Гартман М., Общая биология, пер.(перевод) с нем.(немецкий), 2 изд., М.— Л., 1936; Догель В. А., Зоология беспозвоночных, 6 изд., М., 1975.
А. В. Иванов.
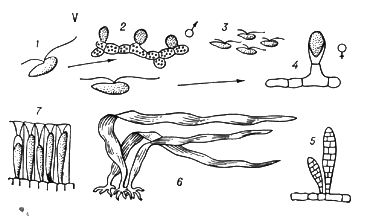
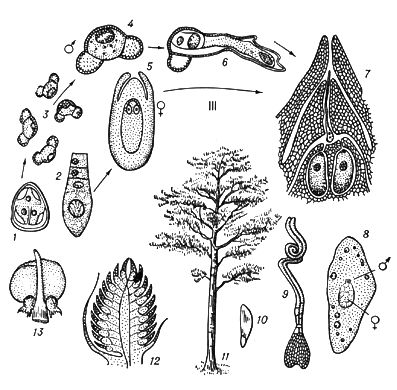
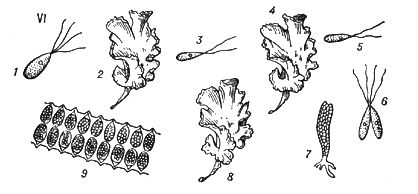
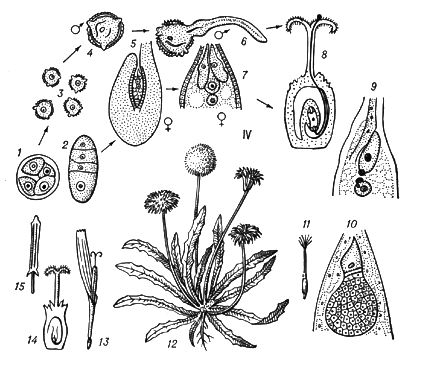
У растений под Ч. п. обычно понимают чередование в циклах развития диплоидной и гаплоидной фаз (рис. 3).
Типичное Ч. п. характерно для растений, у которых многоклеточны как диплоидная фаза (диплонт), так и гаплоидная (гаплонт). Диплонт образует спорангии, в которых в результате мейоза возникают споры (поэтому диплонт наз.(назыв) также спорофитом), а гаплонт — гаметангии, в которых без редукционного деления образуются гаметы (гаплонт наз.(назыв) также гаметофитом); спорофит развивается из зиготы, а гаметофит — из споры. У одних растений (например, у водорослей ульвы, диктиоты) спорофит и гаметофит развиты одинаково, а у др. доминирует либо гаметофит (некоторые бурые водоросли, например кутлерия, все мохообразные), либо спорофит (некоторые бурые водоросли, например ламинария, все папоротникообразные и семенные растения). У многих зелёных и, возможно, некоторых красных водорослей диплоидны только зиготы, делящиеся мейотически, а у сифоновых, диатомовых и некоторых бурых водорослей (как и у огромного большинства животных) гаплоидны только гаметы. У этих растений фактически Ч. п. нет, хотя смена ядерных фаз происходит.
Спорофиты мохообразных, т. н. спорогонии, развиваются на гаметофитах. Гаметофиты папоротникообразных (заростки) существуют самостоятельно, а семенных растений — развиваются на спорофитах. Заростки изоспоровых (равноспоровых) растений обоеполы, гетероспоровых — раздельнополы и более редуцированы (особенно мужские), чем заростки изоспоровых. Так, у покрытосеменных мужской заросток — пыльцевое зерно, женский — зародышевый мешок. См. также Оплодотворение у растений.
Лит.: Тахтаджян А. Л., Высшие растения, т. 1 — От псилофитовых до хвойных, М.—Л., 1956; Поддубная-Арнольди В. А., Цитоэмбриология покрытосеменных растений, М., 1976.