Ядро клеточное, обязательная, наряду с цитоплазмой, составная часть клетки у простейших, многоклеточных животных и растений, содержащая хромосомы и продукты их деятельности. По наличию или отсутствию в клетках Я. все организмы делят на эукариот и прокариот. У последних нет оформленного Я. (отсутствует его оболочка), хотя дезоксирибонуклеиновая кислота (ДНК) имеется. В Я. хранится основная часть наследственной информации клетки; содержащиеся в хромосомах гены играют главную роль в передаче наследственных признаков в ряду клеток и организмов. Я. находится в постоянном и тесном взаимодействии с цитоплазмой; в нём синтезируются молекулы-посредники, переносящие генетическую информацию к центрам белкового синтеза в цитоплазме. Т. о., Я. управляет синтезами всех белков и через них — всеми физиологическими процессами в клетке. Поэтому получаемые экспериментально безъядерные клетки и фрагменты клеток всегда погибают; при пересадке Я. в такие клетки их жизнеспособность восстанавливается. Я. впервые наблюдал чешский учёный Я. Пуркине (1825) в яйцеклетке курицы; в растительных клетках Я. описал английский учёный Р. Броун (1831—33), в животных клетках — немецкий учёный Т. Шванн (1838—39).
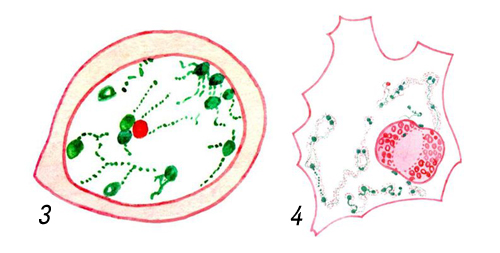
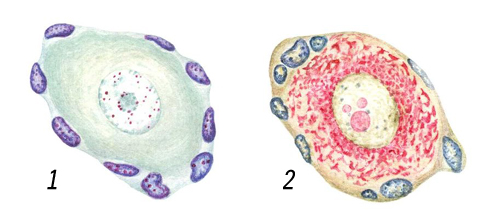
Обычно Я. в клетке одно, находится близ её центра, имеет вид сферического или эллипсоидного пузырька (фигуры 1—3, 5, 6). Реже Я. бывает неправильной (фигура 4) или сложной формы (например, Я. лейкоцитов, макронуклеусы инфузорий). Нередки двух- и многоядерные клетки, обычно образующиеся путём деления Я. без деления цитоплазмы или путём слияния нескольких одноядерных клеток (т. н. симпласты, например поперечнополосатые мышечные волокна). Размеры Я. варьируют от ~ 1 мкм (у некоторых простейших) до ~ 1 мм (некоторые яйцеклетки).
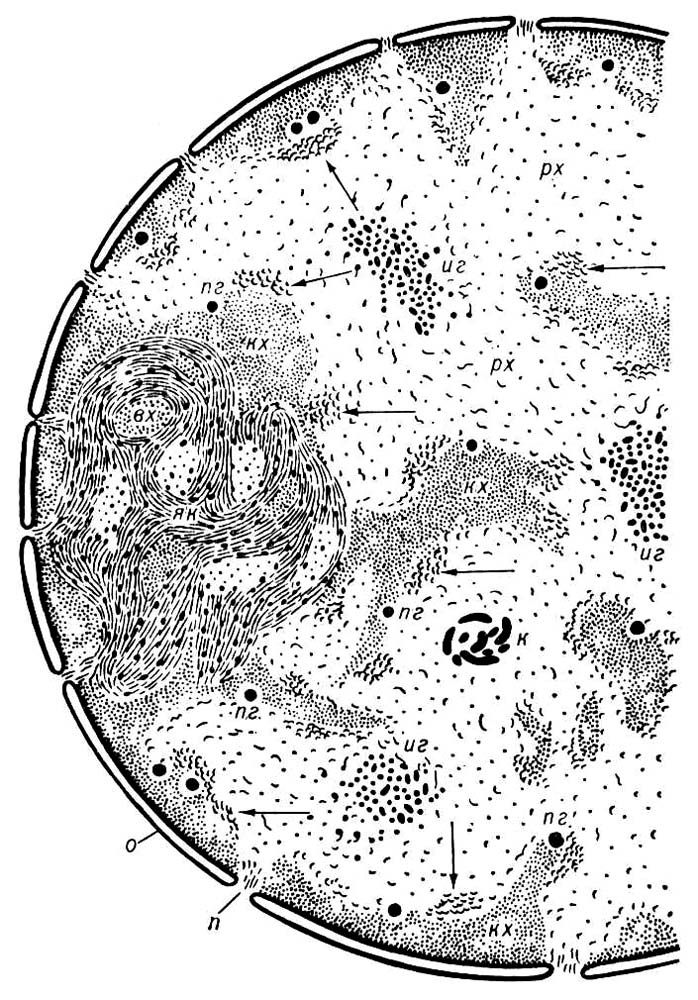
Я. отделено от цитоплазмы ядерной оболочкой (ЯО), состоящей из 2 параллельных липопротеидных мембран толщиной 7—8 нм, между которыми находится узкое перинуклеарное пространство. ЯО пронизана порами диаметром 60—100 нм, на краях которых наружная мембрана ЯО переходит во внутреннюю. Частота пор различна в разных клетках: от единиц до 100—200 на 1 мкм2 поверхности Я. По краю поры располагается кольцо плотного материала — так называемый аннулус. В просвете поры часто имеется центральная гранула диаметром 15—20 нм, соединённая с аннулусом радиальными фибриллами. Вместе с порой эти структуры составляют поровый комплекс, который, по-видимому, регулирует прохождение макромолекул через ЯО (например, вход в Я. белковых молекул, выход из Я. рибонуклеопротеидных частиц и т. п.). Наружная мембрана ЯО местами переходит в мембраны эндоплазматической сети; она обычно несёт белоксинтезирующие частицы — рибосомы. Внутренняя мембрана ЯО иногда образует впячивания в глубь Я. Содержимое Я. представлено ядерным соком (кариолимфой, кариоплазмой) и погруженными в него оформленными элементами — хроматином, ядрышками и др. Хроматин — это более или менее разрыхлённый в неделящемся Я. материал хромосом, комплекс ДНК(дезоксирибонуклеиновая кислота) с белками — так называемый дезоксирибо-нуклеопротеид (ДНП). Он выявляется с помощью цветной реакции Фёльгена на ДНК(дезоксирибонуклеиновая кислота) (фигуры 1 и 8). При делении Я. (см. Митоз) весь хроматин конденсируется в хромосомы; по окончании митоза большая часть участков хромосом опять разрыхляется; эти участки (так называемый эухроматин) содержат в основном уникальные (неповторяющиеся) гены. Другие участки хромосом остаются плотными (так называемый гетерохроматин); в них располагаются главным образом повторяющиеся последовательности ДНК(дезоксирибонуклеиновая кислота). В неделящемся Я. большая часть эухроматина представлена рыхлой сетью фибрилл ДНП толщиной 10 — 30 нм, гетерохроматин — плотными глыбками (хромоцентрами), в которых те же фибриллы плотно упакованы. Часть эухроматина также может переходить в компактное состояние; такой эухроматин считается неактивным в отношении синтеза РНК(рибонуклеиновая кислота). Хромоцентры обычно граничат с ЯО или ядрышком. Есть данные о том, что фибриллы ДНП закреплены на внутренней мембране ЯО.
В неделящемся Я. происходит синтез (репликация) ДНК(дезоксирибонуклеиновая кислота), изучаемый путём регистрации включенных в Я. меченных радиоактивными изотопами предшественников ДНК(дезоксирибонуклеиновая кислота) (обычно тимидина). Показано, что по длине хроматиновых фибрилл имеется множество участков (так называемых репликонов), каждый со своей точкой начала синтеза ДНК(дезоксирибонуклеиновая кислота), от которой репликация распространяется в обе стороны. Вследствие репликации ДНК(дезоксирибонуклеиновая кислота) удваиваются и сами хромосомы.
В хроматине Я. происходит считывание закодированной в ДНК(дезоксирибонуклеиновая кислота) генетической информации путём синтеза на ДНК(дезоксирибонуклеиновая кислота) молекул матричной, или информационной, РНК(рибонуклеиновая кислота) (см. Транскрипция), а также молекул других типов РНК(рибонуклеиновая кислота), участвующих в белковом синтезе. Специальные участки хромосом (и соответственно хроматина) содержат повторяющиеся гены, которые кодируют молекулы рибосомной РНК(рибонуклеиновая кислота); в этих местах Я. формируются богатые рибонуклеопротеидами (РНП) ядрышки, основная функция которых — синтез РНК(рибонуклеиновая кислота), входящей в состав рибосом. Наряду с компонентами ядрышка в Я. есть и другие виды частиц РНК(рибонуклеиновая кислота). К ним относятся перихроматиновые фибриллы толщиной 3—5 нм и перихроматиновые гранулы (ПГ) диаметром 40—50 нм,расположенные на границах зон рыхлого и компактного хроматина. И те и другие, вероятно, содержат матричную РНК(рибонуклеиновая кислота) в соединении с белками, а ПГ отвечают её неактивной форме; наблюдался выход ПГ из Я. в цитоплазму через поры ЯО. Имеются также интерхроматиновые гранулы (20—25 нм), а иногда и толстые (40—60 нм) нити РНП, скрученные в клубки. В ядрах амёб имеются нити РНП, скрученные в спирали (30—35 нм х 300 нм); спирали могут выходить в цитоплазму и, вероятно, содержат матричную РНК(рибонуклеиновая кислота). Наряду с ДНК(дезоксирибонуклеиновая кислота)- и РНК(рибонуклеиновая кислота)-содержащими структурами некоторые Я. содержат чисто белковые включения в виде сфер (например, в Я. растущих яйцеклеток многих животных, в Я. ряда простейших), пучков фибрилл или кристаллоидов (например, в ядрах многих тканевых клеток животных и растений, макронуклеусах ряда инфузорий). В Я. обнаружены также фосфолипиды, липопротепды, ферменты (ДНК-полимераза, РНК(рибонуклеиновая кислота)-полимераза, комплекс ферментов оболочки Я., в том числе аденозинтрифосфатаза, и др.).
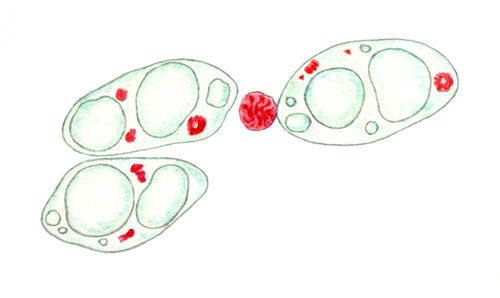
В природе встречаются различные специальные типы Я.: гигантские Я. растущих. яйцеклеток, особенно рыб и земноводных; Я., содержащие гигантские политенные хромосомы (см. Политения), например в клетках слюнных желёз двукрылых насекомых; компактные, лишённые ядрышек Я. сперматозоидов и микронуклеусыинфузорий, сплошь заполненные хроматином и не синтезирующие РНК(рибонуклеиновая кислота); Я., в которых хромосомы постоянно конденсированы, хотя ядрышки образуются (у некоторых простейших, в ряде клеток насекомых); Я., в которых произошло дву- или многократное увеличение числа наборов хромосом (полиплоидия; фигуры 7, 9).
Основной способ деления Я. — митоз, характеризующийся удвоением и конденсацией хромосом, разрушением ЯО (исключение — многие простейшие и грибы) и правильным расхождением сестринских хромосом в дочерние клетки. Однако Я. некоторых специализированных клеток, особенно полиплоидные, могут делиться простой перешнуровкой (см. Амитоз). Высокополиплоидные Я. могут делиться не только на 2, но и на много частей, а также почковаться (фигура 7). При этом может происходить разделение целых хромосомных наборов (т. н. сегрегация геномов).
Лит.: Руководство по цитологии, т. 1, М. —Л., 1965; Райков И. Б., Кариология простейших, Л., 1967; Робертис Э., Новинский В., Саэс Ф.,. Биология клетки, пер.(перевод) с англ.(английский), М., 1973; Ченцов Ю. С., Поляков В. Ю., Ультраструктура клеточного ядра, М., 1974; The nucleus, ed. A. J. Dalton, F, Haguenau, N. Y. — L., 1968; The cell nucleus, ed. Н. Busch, v. 1—3, N. Y. — L., 1974.