Віруси (від латів.(латинський) Virus — отрута), віруси, що фільтруються, ультравіруси, збудники інфекційних хвороб рослин, тварин і людини, що розмножуються лише в живих клітинах. Ст дрібніше за більшість відомих мікробів; майже всі Ст проходят через бактерійні фільтри . На відміну від бактерій, Ст не удається культивувати на звичайних живильних середовищах. Для експериментальних і медичних цілей (здобуття вакцин і ін.) Ст культивують в тваринах і рослинних організмах, курячих ембріонах і в культурах тканин і кліток. Ст викликають багато захворювань: віспу, кір, грип, поліомієліт, чуму рогатої худоби і птиць, сказ, ряд захворювань риб і земноводних, жовтяницю шовкопряда, мозаїчну хворобу тютюну закуклювання вівса, багато захворювань грибів і синьо-зелених водоростей і ін. (див. Вірусні хвороби, Вірусні хвороби рослин ). Обширний загін Ст, що приголомшують бактерії, складають бактеріофаги .
Існування тих, що проходять через бактерійних фільтри збудників інфекційних хвороб було вперше показано в 1892 Д. І. Івановським, що відкрив фільтрованість збудника мозаїчної хвороби тютюну. Незабаром була доведена фільтрованість збудників ящура (1897), чуми рогатої худоби (1899), віспи птиць (1902), сказу (1903) і ін. У сучасному сенсі слово «В.» вперше застосував М. Бейерінк (1899); раніше Ст інколи називали і хвороботворні мікроби, наприклад збудника туберкульозу. У міру вивчення Ст усе більш уточнюється і звужується поняття про них. Збудники ряду хвороб, відношувані раніше до Ст, наприклад рикетсії і збудники пситтакоза, виключені з цієї групи організмів. Зрілі частки Ст — віріони, або віроспори пристосовані до перенесення несприятливих умов поза організмом і не виявляють на цій стадії жодних ознак життя. Попавши в організм, в чутливих к В. клітки, віроспори переходять в стадію розвитку і розмноження, яка завершується утворенням дочірніх зрілих часток Ст
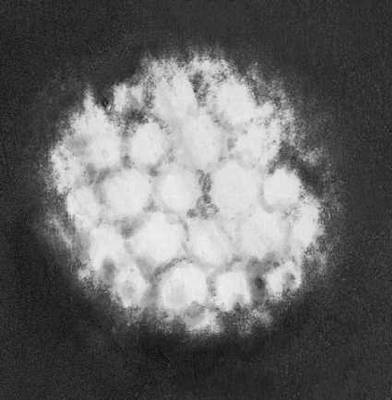
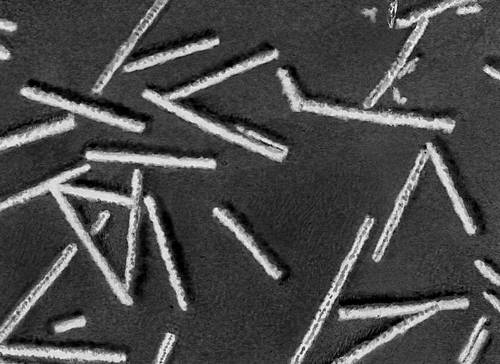
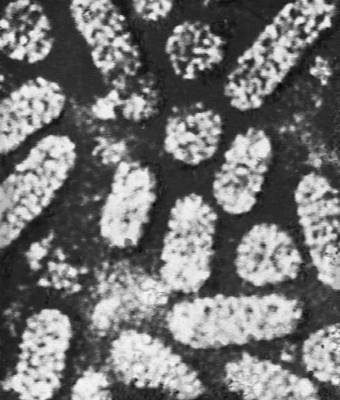
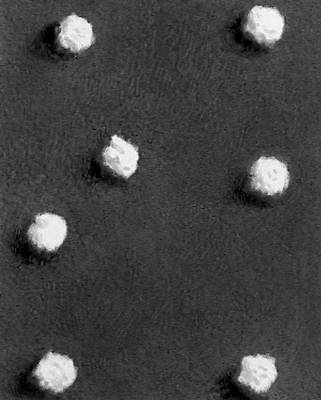
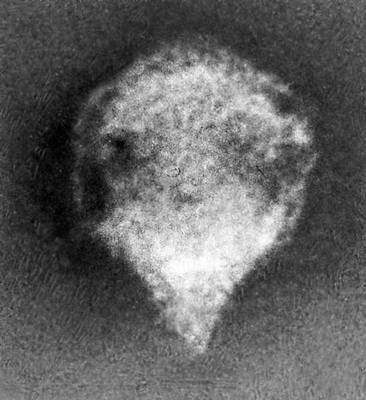
Будова і склад часток В. Форма віріонів дуже всіляка. В багатьох бактеріофагів вони складаються з голівки і відростка, в Ст віспи вони прямокутні, в Ст герпесу і грипу — кулясті, в Ст мозаїчної хвороби тютюну — палочкообразниє, в Ст мозаїчної хвороби картоплі — ниткоподібні, в Ст поліомієліту і жовтої мозаїки турнепсу — багатогранні кульки, в Ст сказу, а також мозаїки пшениці і люцерни — контурами схожі на палички бактерій або нагадують кулю. По розмірах Ст ділять на великих (300—400 нм в діаметрі), середніх (80—125 нм ) і дрібних (20—30 нм ). Крупні Ст можна бачити в світловий мікроскоп (звичайний, фазово-контрастний, люмінесцентний); останні вивчають лише за допомогою електронного мікроскопа. Дані про розміри часток Ст отримані методами ультрафільтрації, фракційного і аналітичного ультрацентрофугування, електрофорезу в гелях і електронній мікроскопії (таблиця.).
Розміри деяких вірусів
(для порівняння дані розміри еритроцита, бактерії і деяких молекул)*
Об'єкт ісследо- ванія
Маса (10 6- ат. м. во- дорода)
Діаметр ілі- довжина, умно-
женная на- ширину ( нм )
Еритроцит . . . . . . . .
173000000
7500
Кишкова палоч- но . . . . . . . . . . . . . . .
180000
(1000—3000) 5000
Вірус вакцини . . . .
2300
262∙209
« герпесу . . . . . .
1400
213∙175
« гріїппа . . . . . .
700
103∙90
« бактерії Т 2 . . . .
120
80∙60
(голівка)
100∙20
(хвіст)
« мозаїчною . . .
хвороби та- бака . . . . . . . .
39,2
300∙15
« Х картофе- ля . . . . . . . . . .
39,0
(500—580)∙10
« поліоміє- льоту . . . . . . . .
6,7
28
« жовтої мо- заїки тур- непса . . . . . . .
5,1
28
« ящура . . . . . .
5,1
28
Білок гемоціанин . .
6,7
59∙13,2
« гемоглобіну
коня . . . . .
0,069
2,8∙0,6
« курячого
яйця . . . . . . . .
0,040
1,8∙0,6
* Різні автори залежно від вживаних ними методів і ін. умов отримували величини, що відхиляються від тих, що приводяться, проте порядок величин у всіх випадках зберігається.
В будові різних віріонів є багато загального. Всі вони мають білкову оболонку — капсид і внутрішній вміст — нуклеокапсид, що складається головним чином з нуклеїнової кислоти (НК) — ДНК(дезоксирибонуклеїнова кислота) або РНК(рибонуклеїнова кислота). Багато Ст мають поверхневу оболонку, покриваючу білкову. Окремі елементи білкової оболонки називаються капсомерами. В деяких Ст (наприклад, мозаїчній хворобі тютюну) НК у вигляді спіралі включена в білкову оболонку, без руйнування якої не може бути звільнена. В інших Ст (наприклад, жовтої мозаїки турнепсу) спіральний закручена нитка НК лежить в капсиді, як в коробочці, і може вийти звідти без руйнування оболонки. НК — носії спадкової інформації про будову і властивості В.; білки Ст захищають НК, а також обумовлюють ферментативні і антигенні властивості Ст (див. Антигени, Ферменти ). Будова вірусних часток, пристосованих до перенесення несприятливих умов, може бути і складнішим; такі, наприклад, поліедри, що утворюються деякими Ст комах (вони складаються з оболонки, кристалічної білкової маси і включених в неї часток Ст).
Хімічний склад різних Ст неоднаковий. Одні Ст містять ліпіди; серед них є Ст з ДНК(дезоксирибонуклеїнова кислота) (віспи, герпесу і ін.), з РНК(рибонуклеїнова кислота)(грипу, пташиної чуми, саркоми Раусу, бронзовості помідора, жовтої карликовості картоплі і ін.). В інших. Ст ліпіди відсутні. У цій групі також є Ст з ДНК(дезоксирибонуклеїнова кислота) (аденовіруси, більшість бактеріофагів, Ст жовтяниці шовкопряда) і з РНК(рибонуклеїнова кислота) (поліомієліту, ящура; більшість Ст, що викликають хвороби рослин; деякі бактеріофаги). Окрім ліпідів, білка і нуклеїнової кислоти, в Ст зустрічаються в невеликій кількості поліаміни (путресцин, спермідін і ін.), інколи вітаміни (вітамін B 2 фолієвая кислота), а також ряд металів; у деяких Ст містяться з'єднання білка з полісахаридами.
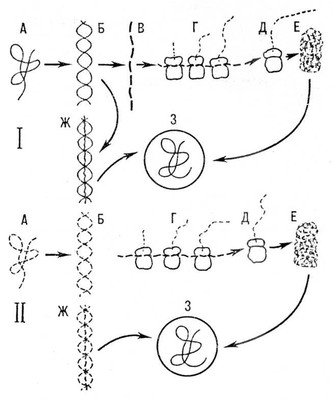
Розмноження Ст відбувається в клітках. Бактеріофаги розчиняють оболонку бактерії і вводять в бактерію нитку НК, причому капсид фага залишається поза кліткою. Багато Ст поглинаються кліткою дорогою піноцитозу . Попавши в клітку, вони звільняються від оболонки. Перші етапи розвитку Ст в клітці в загальних межах полягають в тому, що будуються так звані ранні білки, тобто білки-ферменти, необхідні Ст для реплікації (подвоєння) їх НК. Так звані пізні білки беруть участь в утворенні білкових оболонок дочірніх віроспор. З ферментів в Ст, що містять ДНК(дезоксирибонуклеїнова кислота), одним з перших синтезується полімераза РНК(рибонуклеїнова кислота), яка будує на нитці ДНК(дезоксирибонуклеїнова кислота) інформаційну РНК(рибонуклеїнова кислота) (І-РНК). Ета РНК(рибонуклеїнова кислота) потрапляє на рибосоми клітки, де і відбувається синтез інших білків вірусної частки (див. Білки, розділ Біосинтез). Ст, що містять РНК(рибонуклеїнова кислота), синтезують полімеразу, що каталізує синтез нових часток вірусною РНК(рибонуклеїнова кислота); ця РНК(рибонуклеїнова кислота) переходить на рибосоми і контролює синтез білка капсиду. Таким чином, Ст, що містять РНК(рибонуклеїнова кислота), не потребують ДНК(дезоксирибонуклеїнова кислота) для розмноження і передачі генетичної інформації потомству (см. схему ).
Від цієї загальної схеми розмноження Ст є різні відхилення. Так, деякі Ст містять білки-ферменти; Ст осповакцини синтезує в клітці господаря подвійні нитки РНК(рибонуклеїнова кислота) і т.д. Багато особливостей розмноження Ст ще не з'ясовано. Існують, наприклад, особливі вогнища розмноження ниток НК, і при дозріванні часток Ст синтезується білок, що охоплює окремі відрізки НК. Інколи цей процес йде недосконало, утворюються неповноцінні частки Ст, в яких немає або мало вмісту, це — так звані неінфекційні В. Во багатьох випадках вогнища розмноження Ст добре видно в клітці під мікроскопом. Ці вогнища називаються внутріклітинними включеннями, або Х-тіламі. Коли Х-тіло закінчує свій розвиток, в нім утворюється віроспора. В багатьох Ст віроспори утворюють в Х-тілах кристалічні агрегати, в інших Ст вони невідомі. Деякі Ст розмножуються в ядрі клітки інші — в її цитоплазмі, треті — і в ядрі, і в цитоплазмі. НК знаходиться у віроспоре в спеціально закрученому стані. Довжина нитки НК в різних Ст різна. Так, в Ст віспи вона досягає 83 мкм , в крупних бактеріофагів, наприклад Т 4 , — 70 мкм . В найдрібніших бактеріофагів нитка НК має довжину близько 2 мкм . Залежно від довжини нитки НК (що визначає об'єм спадкової інформації, яку має в своєму розпорядженні те або інше Ст), тобто від здатності Ст синтезувати більш менш всілякі молекули білків, різна міра участі складових частин клітки-господаря в розмноженні Ст і їх побудові. Ст, що мають нитку НК значної довжини, можуть синтезувати багато речовин. Так, деякі бактеріофаги синтезують в клітці декілька десятків різних білків. Всі Ст, що містять ДНК(дезоксирибонуклеїнова кислота), синтезують власну РНК(рибонуклеїнова кислота). Навіть якщо клітка-господар має необхідні для Ст ферменти, Ст дуже часто синтезують власні ферменти, що володіють подібною дією. Найдрібніші фаги володіють інформацією для синтезу лише трьох власних білків; наприклад, фаг МЗ-2 синтезує залежну від РНК(рибонуклеїнова кислота) полімеразу і два білки, необхідні для побудови зрілих часток В. Таким образом, міра залежності Ст від різних ферментів клітки-господаря різна. Деякі Ст такі бідні спадковою (генетичною) інформацією, що можуть розмножуватися в клітці лише у присутності ін. В. Завісимость Ст не лише від клітки, але і від інших Ст існує, наприклад, між Ст некрозу тютюну і його супутником, віроспори якого дрібніше за віроспор некроз тютюну. Ще тісніші взаємини існують між деякими Ст, що приголомшують тварин і людину. Серед Ст, здатних викликати злоякісні пухлини (див. Опухолеродниє віруси ), відомі Ст з дефектною часткою, яка не може утворювати власну білкову оболонку. Ці Ст досягають зрілого стану, лише якщо вони розмножуються у присутності інших Ст (такі стосунки, наприклад, між опухолеродним мавпячим вірусом S-40 і деякими аденовірусами ). НК опухолеродного Ст в цьому випадку включається в капсид аденовіруса і разом з ним потрапляє в чутливу клітку. Вихід Ст з клітки в одних випадках здійснюється лише при руйнуванні клітки (багато фаги, Ст віспи), в інших — частки Ст покидають клітку, не вбиваючи її при цьому (міксовіруси, деякі дрібні фаги).
Якщо в клітку потрапляють Ст, що розрізняються по тих же або інших генам (відмінність може бути результатом мутації ), то в потомстві можна спостерігати Ст, що сполучають властивості два і більше вихідних форм. Це вказує на наявність обміну (перекомбінациі) ознак таких форм при розмноженні Ст в одній клітці. Закономірності цих процесів вивчає генетика Ст (див. Генетика мікроорганізмів ).
Стійкість віроспор до зовнішніх дій різна, але здебільше велика. Деякі Ст інактівіруются лише при нагріванні до 90°С (Ст мозаїчної хвороби тютюну), легко переносять дуже низькі температури (—70°С і нижче), а також висушування.
Способи поширення Ст в природі різні: багато хто з них може безпосередньо заражати чутливий організм (Ст грипу, віспи, мозаїчної хвороби тютюну, бактеріофаги), інші циркулюють в природі складнішим чином і переносяться за допомогою інших організмів. Так, Ст некрозу тютюну передається за допомогою грибка, що мешкає в грунті (Olpidium): останній, проникаючи в коріння рослини, вносить і В. Багато Ст передаються паразитуючими в рослин нематодами . Ст тварин, людини і рослин переносять також кліщі і комахи. Передача одних Ст членистоногими, що смокчуть, носить механічний характер; у інших випадках Ст проробляють частину свого розвитку в переноснику і навіть можуть передаватися з яйцями переносника з покоління в покоління. Багато Ст, що приголомшують людину і домашніх тварин, мешкають і в диких тваринах; приголомшуючі культурні рослини — в диких рослинах і бур'янах.
Спроби виявити життєдіяльність віроспор поза кліткою, природно, не увінчалися успіхом: відомо, що форми життя, що покояться, взагалі не виявляють життєдіяльності (див. Анабіоз ). У безклітинних системах можна відтворити окремі етапи розмноження Ст, отримати саморепродукцию вірусної НК, а також під контролем цієї НК — синтез білків, характерних для В. Но ці процеси йдуть лише у присутності тих, що витягують з клітки рибосом; отже, ці системи, хоча і є безклітинними, не можуть розглядатися як що сповна штучно синтезуються.
Про походження Ст є різні припущення. Деякі вважають, що Ст можуть спонтанно зароджуватися в організмі господаря під впливом несприятливих умов. Але ця думка спростовується слідами тривалої еволюції Ст (їх пристосування до циркуляції в природі), а також відсутністю перехідних форм між Ст і органоїдами клітки. Інші дослідники думають, що Ст — нащадки простих форм життя, проте і це припущення маловірогідне, оскільки виражений паразитичний характер Ст передбачає існування більш високоорганізованих істот, в яких Ст могли б жити і розмножуватися. Тому найймовірніше, що Ст виникли від що вільно живуть більш за складно організовані форми, і простота Ст вторинна, вона — результат пристосування до паразитичного способу життя. Така вторинна простота, пов'язана з втратою пристосованості до самостійного живлення і посиленням здібності до розмноження, взагалі дуже характерна для паразитів. На користь старовини Ст і тривалої їх еволюції говорить також те, що вони вступають в складні взаємини з іншими видами тварин і рослин (трансмісивні Ст, передавані різними тваринами).
Систематика В. Общепрінятой класифікації і позначення Ст ще немає. Їм дають, як і іншим тваринам і рослинам, родові і видові назви, користуються народними позначеннями, різними скороченнями або ставлять родову назву організму, приголомшуваного Ст, і номер (наприклад, Nicotiana virus I — Ст мозаїчної хвороби тютюну). Тому кожне Ст може мати декілька назв. Першу спробу систематики Ст зробив чеський учений Г. Провачек (1907); він відніс Ст до тварин, до групи Chamydozoa. До середини 20 ст склалося 3 головних напряму в систематиці Ст Прибічники одного в основу системи Ст кладуть властивості віріонів; при цьому враховують присутність в них РНК(рибонуклеїнова кислота) або ДНК(дезоксирибонуклеїнова кислота), симетрію нуклеокапсиду, наявність або відсутність пеплоса (особливої оболонки капсиду), діаметр нуклеокапсиду (в спіральних віріонів), число граней і капсомерів (в кубічних віріонів). Представники другого напряму (нумерічеськая система), враховуючи по можливості всі ознаки, об'єднують ті Ст, в яких більше загальних ознак. Прибічники третього напряму, зберігаючи принципи класичної систематики, об'єднують Ст в групи на основі істотних ознак, що характеризують їх спорідненість (хімічна близькості, схожість морфологічних стадій розвитку і способів циркуляції в природі). Міжнародний комітет з номенклатури Ст пропонує користуватися бінарною номенклатурою, додаючи до родової назви слово «В.» (наприклад, рід Ст віспи — Poxvirus). Багато загальноприйнятих назв зберігають, хоча вони і не відповідають бінарній номенклатурі. Прибічники нумерічеськой системи пропонують користуватися криптограмами, які в умовних позначеннях розшифровують найважливіші властивості Ст Так, Ст тютюнової мозаїки позначають так: Перший член показує, що це Ст містить РНК(рибонуклеїнова кислота) (R) і вона в нім однониткова (1); другий член — молекулярну масу РНК(рибонуклеїнова кислота) в мільйонах і % РНК(рибонуклеїнова кислота) в частці; третій — що форма цієї частки подовжена з паралельними сторонами і кінці не закруглені, а також що подібну форму має і нуклеокапсид; останній член вказує, що вірус приголомшує вищі рослини (S) і поширюється без переносника (0).
Літ.: Рижков Ст Л., Віруси, в сб.(збірка): Очима ученого, М., 1963; Стенлі В., Веленс Е., Віруси і природа життя, пер.(переведення) з англ.(англійський), М., 1963; Вірусологія і імунологія, під ред. Л. А. Зільбера, М., 1964; Молекулярні основи біології вірусів, М., 1966; Стент Р., Молекулярна біологія вірусів бактерій, пер.(переведення) з англ.(англійський), М., 1965; Ендрюс До., Природна історія вірусів, пер.(переведення) з англ.(англійський), М., 1969; The viruses, ed. F. М. Burnet and W. М. Stanley, v. 1—3, N. Y. — L., 1959; Fenner F. J., The biology of animal viruses, v. 1, N. Y., 1968; Gibbs A., Plant virus classification, «Advances in Virus Research», 1969, v. 14, р. 263—328.