Вирусы (от лат.(латинский) Virus — яд), фильтрующиеся вирусы, ультравирусы, возбудители инфекционных болезней растений, животных и человека, размножающиеся только в живых клетках. В. мельче большинства известных микробов; почти все В. проходят через бактериальные фильтры. В отличие от бактерий, В. не удаётся культивировать на обычных питательных средах. Для экспериментальных и медицинских целей (получения вакцини др.) В. культивируют в животных и растительных организмах, куриных эмбрионах и в культурах тканей и клеток. В. вызывают многие заболевания: оспу, корь, грипп, полиомиелит, чуму рогатого скота и птиц, бешенство, ряд заболеваний рыб и земноводных, желтуху шелкопряда, мозаичную болезнь табака, закукливание овса, многие заболевания грибов и сине-зелёных водорослей и др. (см. Вирусные болезни, Вирусные болезни растений). Обширный отряд В., поражающих бактерии, составляют бактериофаги.
Существование проходящих через бактериальные фильтры возбудителей инфекционных болезней было впервые показано в 1892 Д. И. Ивановским, открывшим фильтруемость возбудителя мозаичной болезни табака. Вскоре была доказана фильтруемость возбудителей ящура (1897), чумы рогатого скота (1899), оспы птиц (1902), бешенства (1903) и др. В современном смысле слово «В.» впервые применил М. Бейеринк (1899); раньше В. иногда называли и болезнетворных микробов, например возбудителя туберкулёза. По мере изучения В. всё более уточняется и суживается понятие о них. Возбудители ряда болезней, относимые ранее к В., например риккетсии и возбудители пситтакоза, исключены из этой группы организмов. Зрелые частицы В. — вирионы, или вироспоры, приспособлены к перенесению неблагоприятных условий вне организма и не обнаруживают на этой стадии никаких признаков жизни. Попав в организм, в чувствительные к В. клетки, вироспоры переходят в стадию развития и размножения, которая завершается образованием дочерних зрелых частиц В.
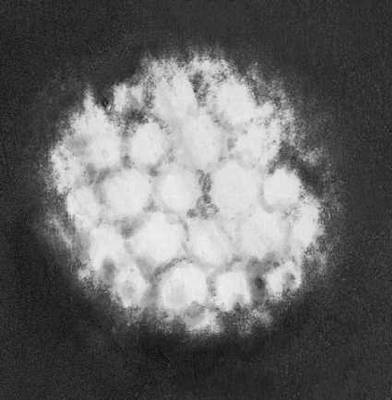
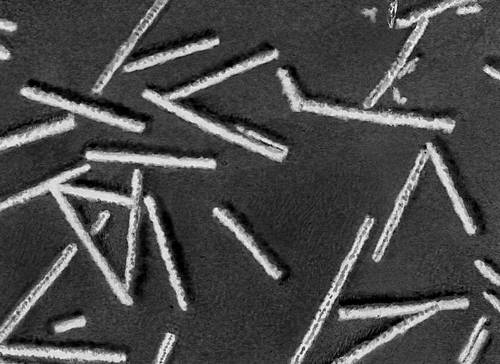
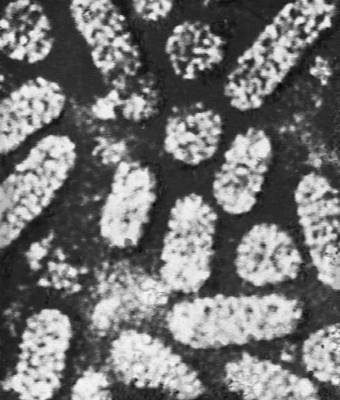
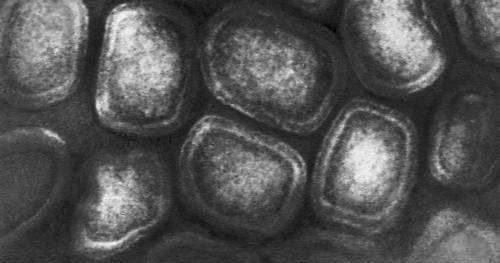
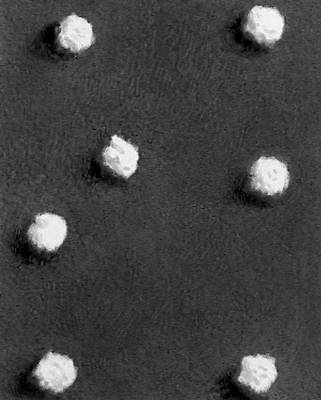
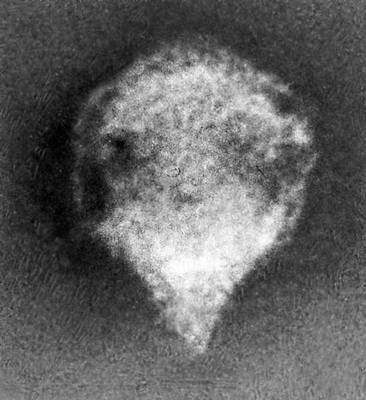
Строение и состав частиц В. Форма вирионов очень разнообразна. У многих бактериофагов они состоят из головки и отростка, у В. оспы они прямоугольные, у В. герпеса и гриппа — шарообразные, у В. мозаичной болезни табака — палочкообразные, у В. мозаичной болезни картофеля — нитевидные, у В. полиомиелита и жёлтой мозаики турнепса — многогранные шарики, у В. бешенства, а также мозаики пшеницы и люцерны — очертаниями похожи на палочки бактерий или напоминают пулю. По размерам В. делят на крупные (300—400 нм в диаметре), средние (80—125 нм) и мелкие (20—30 нм). Крупные В. можно видеть в световой микроскоп (обычный, фазово-контрастный, люминесцентный); остальные изучают только с помощью электронного микроскопа. Данные о размерах частиц В. получены методами ультрафильтрации, фракционного и аналитического ультрацентрифугирования, электрофореза в гелях и электронной микроскопии (табл.).
Размеры некоторых вирусов
(для сравнения даны размеры эритроцита, бактерии и некоторых молекул)*
Объект исследо- вания
Масса (106- ат. м. во- дорода)
Диаметр или- длина, умно-
женная на- ширину (нм)
Эритроцит . . . . . . . .
173000000
7500
Кишечная палоч- ка . . . . . . . . . . . . . . .
180000
(1000—3000) 5000
Вирус вакцины . . . .
2300
262∙209
« герпеса . . . . . .
1400
213∙175
« грииппа . . . . . .
700
103∙90
« бактерии Т2 . . . .
120
80∙60
(головка)
100∙20
(хвост)
« мозаичной . . .
болезни та- бака . . . . . . . .
39,2
300∙15
« Х картофе- ля . . . . . . . . . .
39,0
(500—580)∙10
« полиомие- лита . . . . . . . .
6,7
28
« жёлтой мо- заики тур- непса . . . . . . .
5,1
28
« ящура . . . . . .
5,1
28
Белок гемоцианин . .
6,7
59∙13,2
« гемоглобина
лошади . . . . .
0,069
2,8∙0,6
« куриного
яйца . . . . . . . .
0,040
1,8∙0,6
* Разные авторы в зависимости от применяемых ими методов и др. условий получали величины, отклоняющиеся от приводимых, однако порядок величин во всех случаях сохраняется.
В строении разных вирионов есть много общего. Все они имеют белковую оболочку — капсид и внутреннее содержимое — нуклеокапсид, состоящее главным образом из нуклеиновой кислоты(НК) — ДНК(дезоксирибонуклеиновая кислота) или РНК(рибонуклеиновая кислота). Многие В. имеют поверхностную оболочку, покрывающую белковую. Отдельные элементы белковой оболочки называются капсомерами. У некоторых В. (например, мозаичной болезни табака) НК в виде спирали включена в белковую оболочку, без разрушения которой не может быть освобождена. У других В. (например, жёлтой мозаики турнепса) спирально закрученная нить НК лежит в капсиде, как в коробочке, и может выйти оттуда без разрушения оболочки. НК — носители наследственной информации о строении и свойствах В.; белки В. защищают НК, а также обусловливают ферментативные и антигенные свойства В. (см. Антигены, Ферменты). Строение вирусных частиц, приспособленных к перенесению неблагоприятных условий, может быть и более сложным; таковы, например, полиэдры, образуемые некоторыми В. насекомых (они состоят из оболочки, кристаллической белковой массы и включенных в неё частиц В.).
Химический состав разных В. неодинаков. Одни В. содержат липиды; среди них есть В. с ДНК(дезоксирибонуклеиновая кислота) (оспы, герпеса и др.), с РНК(рибонуклеиновая кислота) (гриппа, птичьей чумы, саркомы Рауса, бронзовости помидора, жёлтой карликовости картофеля и др.). У других. В. липиды отсутствуют. В этой группе также есть В. с ДНК(дезоксирибонуклеиновая кислота) (аденовирусы, большинство бактериофагов, В. желтухи шелкопряда) и с РНК(рибонуклеиновая кислота) (полиомиелита, ящура; большинство В., вызывающих болезни растений; некоторые бактериофаги). Кроме липидов, белка и нуклеиновой кислоты, в В. встречаются в небольшом количестве полиамины (путресцин, спермидин и др.), иногда витамины (витамин B2, фолиевая кислота), а также ряд металлов; в некоторых В. содержатся соединения белка с полисахаридами.
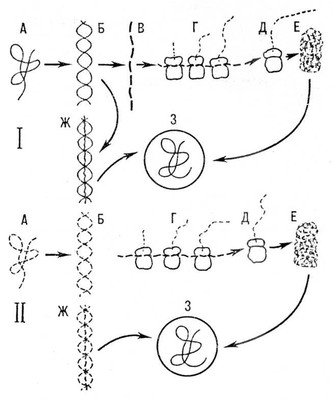
Размножение В. происходит в клетках. Бактериофаги растворяют оболочку бактерии и вводят в бактерию нить НК, причём капсид фага остаётся вне клетки. Многие В. поглощаются клеткой путём пиноцитоза. Попав в клетку, они освобождаются от оболочки. Первые этапы развития В. в клетке в общих чертах состоят в том, что строятся так называемые ранние белки, т. е. белки-ферменты, необходимые В. для репликации (удвоения) их НК. Так называемые поздние белки участвуют в образовании белковых оболочек дочерних вироспор. Из ферментов у В., содержащих ДНК(дезоксирибонуклеиновая кислота), одним из первых синтезируется полимераза РНК(рибонуклеиновая кислота), которая строит на нити ДНК(дезоксирибонуклеиновая кислота) информационную РНК(рибонуклеиновая кислота) (и-РНК). Эта РНК(рибонуклеиновая кислота) попадает на рибосомы клетки, где и происходит синтез других белков вирусной частицы (см. Белки, раздел Биосинтез). В., содержащие РНК(рибонуклеиновая кислота), синтезируют полимеразу, катализирующую синтез новых частиц вирусной РНК(рибонуклеиновая кислота); эта РНК(рибонуклеиновая кислота) переходит на рибосомы и контролирует синтез белка капсида. Таким образом, В., содержащие РНК(рибонуклеиновая кислота), не нуждаются в ДНК(дезоксирибонуклеиновая кислота) для размножения и передачи генетической информации потомству (см. схему).
От этой общей схемы размножения В. имеются различные отклонения. Так, некоторые В. содержат белки-ферменты; В. осповакцины синтезирует в клетке хозяина двойные нити РНК(рибонуклеиновая кислота) и т.д. Многие особенности размножения В. ещё не выяснены. Существуют, например, особые очаги размножения нитей НК, и при созревании частиц В. синтезируется белок, охватывающий отдельные отрезки НК. Иногда этот процесс идет несовершенно, образуются неполноценные частицы В., в которых нет или мало содержимого, это — так называемые неинфекционные В. Во многих случаях очаги размножения В. хорошо видны в клетке под микроскопом. Эти очаги называются внутриклеточными включениями, или Х-телами. Когда Х-тело заканчивает свое развитие, в нём образуется вироспора. У многих В. вироспоры образуют в Х-телах кристаллические агрегаты, у других В. они неизвестны. Некоторые В. размножаются в ядре клетки другие — в ее цитоплазме, третьи — и в ядре, и в цитоплазме. НК находится в вироспоре в специально закрученном состоянии. Длина нити НК у разных В. различна. Так, у В. оспы она достигает 83 мкм, у крупных бактериофагов, например Т4, — 70 мкм. У мельчайших бактериофагов нить НК имеет длину около 2 мкм. В зависимости от длины нити НК (что определяет объем наследственной информации, которой располагает тот или иной В.), т. е. от способности В. синтезировать более или менее разнообразные молекулы белков, различна степень участия составных частей клетки-хозяина в размножении В. и их построении. В., имеющие нить НК значительной длины, могут синтезировать многие вещества. Так, некоторые бактериофаги синтезируют в клетке несколько десятков разных белков. Все В., содержащие ДНК(дезоксирибонуклеиновая кислота), синтезируют собственную РНК(рибонуклеиновая кислота). Даже если клетка-хозяин имеет необходимые для В. ферменты, В. очень часто синтезируют собственные ферменты, обладающие подобным действием. Мельчайшие фаги обладают информацией для синтеза только трёх собственных белков; например, фаг МЗ-2 синтезирует зависящую от РНК(рибонуклеиновая кислота) полимеразу и два белка, необходимые для построения зрелых частиц В. Таким образом, степень зависимости В. от различных ферментов клетки-хозяина различна. Некоторые В. так бедны наследственной (генетической) информацией, что могут размножаться в клетке только в присутствии др. В. Зависимость В. не только от клетки, но и от других В. существует, например, между В. некроза табака и его спутником, вироспоры которого мельче вироспор некроза табака. Ещё более тесные взаимоотношения существуют между некоторыми В., поражающими животных и человека. Среди В., способных вызывать злокачественные опухоли (см. Опухолеродные вирусы), известны В. с дефектной частицей, которая не может образовывать собственную белковую оболочку. Эти В. достигают зрелого состояния, только если они размножаются в присутствии других В. (таковы отношения, например, между опухолеродным обезьяньим вирусом S-40 и некоторыми аденовирусами). НК опухолеродного В. в этом случае включается в капсид аденовируса и вместе с ним попадает в чувствительную клетку. Выход В. из клетки в одних случаях совершается только при разрушении клетки (многие фаги, В. оспы), в других — частицы В. покидают клетку, не убивая её при этом (миксовирусы, некоторые мелкие фаги).
Если в клетку попадают В., различающиеся по тем же или другим генам (различие может быть результатом мутации), то в потомстве можно наблюдать В., соединяющие свойства двух и больше исходных форм. Это указывает на наличие обмена (перекомбинации) признаков таких форм при размножении В. в одной клетке. Закономерности этих процессов изучает генетика В. (см. Генетика микроорганизмов).
Устойчивость вироспор к внешним воздействиям различна, но по большей части велика. Некоторые В. инактивируются только при нагревании до 90°С (В. мозаичной болезни табака), легко переносят очень низкие температуры (—70°С и ниже), а также высушивание.
Способы распространения В. в природе различны: многие из них могут непосредственно заражать чувствительный организм (В. гриппа, оспы, мозаичной болезни табака, бактериофаги), иные циркулируют в природе более сложным образом и переносятся при помощи других организмов. Так, В. некроза табака передаётся при помощи обитающего в почве грибка (Olpidium): последний, проникая в корни растения, вносит и В. Многие В. передаются паразитирующими у растений нематодами. В. животных, человека и растений переносят также клещи и насекомые. Передача одних В. сосущими членистоногими носит механический характер; в других случаях В. проделывают часть своего развития в переносчике и даже могут передаваться с яйцами переносчика из поколения в поколение. Многие В., поражающие человека и домашних животных, обитают и в диких животных; поражающие культурные растения — в диких растениях и сорняках.
Попытки обнаружить жизнедеятельность вироспор вне клетки, естественно, не увенчались успехом: известно, что покоящиеся формы жизни вообще не обнаруживают жизнедеятельности (см. Анабиоз). В бесклеточных системах можно воспроизвести отдельные этапы размножения В., получить саморепродукцию вирусной НК, а также под контролем этой НК — синтез белков, характерных для В. Но эти процессы идут только в присутствии извлечённых из клетки рибосом; следовательно, эти системы, хотя и являются бесклеточными, не могут рассматриваться как вполне искусственно синтезируемые.
О происхождении В. имеются различные предположения. Некоторые считают, что В. могут спонтанно зарождаться в организме хозяина под влиянием неблагоприятных условий. Но это мнение опровергается следами длительной эволюции В. (их приспособление к циркуляции в природе), а также отсутствием переходных форм между В. и органоидами клетки. Другие исследователи думают, что В. — потомки простейших форм жизни, однако и это предположение маловероятно, так как выраженный паразитический характер В. предполагает существование более высокоорганизованных существ, в которых В. могли бы жить и размножаться. Поэтому наиболее вероятно, что В. возникли от свободно живущих более сложно организованных форм, и простота В. вторична, она — результат приспособления к паразитическому образу жизни. Такая вторичная простота, связанная с утратой приспособленности к самостоятельному питанию и усилением способности к размножению, вообще очень характерна для паразитов. В пользу древности В. и длительной их эволюции говорит также то, что они вступают в сложные взаимоотношения с другими видами животных и растений (трансмиссивные В., передаваемые различными животными).
Систематика В. Общепринятой классификации и обозначения В. ещё нет. Им дают, как и другим животным и растениям, родовые и видовые названия, пользуются народными обозначениями, различными сокращениями или ставят родовое название организма, поражаемого В., и номер (например, Nicotiana virus I — В. мозаичной болезни табака). Поэтому каждый В. может иметь несколько названий. Первую попытку систематики В. сделал чешский учёный Г. Провачек (1907); он отнёс В. к животным, к группе Chamydozoa. К середине 20 в. сложилось 3 главных направления в систематике В. Сторонники одного в основу системы В. кладут свойства вирионов; при этом учитывают присутствие в них РНК(рибонуклеиновая кислота) или ДНК(дезоксирибонуклеиновая кислота), симметрию нуклеокапсида, наличие или отсутствие пеплоса (особой оболочки капсида), диаметр нуклеокапсида (у спиральных вирионов), число граней и капсомеров (у кубических вирионов). Представители второго направления (нумерическая система), учитывая по возможности все признаки, объединяют те В., у которых больше общих признаков. Сторонники третьего направления, сохраняя принципы классической систематики, объединяют В. в группы на основе существенных признаков, характеризующих их родство (химическая близости, сходства морфологических стадий развития и способов циркуляции в природе). Международный комитет по номенклатуре В. предлагает пользоваться бинарной номенклатурой, добавляя к родовому названию слово «В.» (например, род В. оспы — Poxvirus). Многие общепринятые названия сохраняют, хотя они и не соответствуют бинарной номенклатуре. Сторонники нумерической системы предлагают пользоваться криптограммами, которые в условных обозначениях расшифровывают важнейшие свойства В. Так, В. табачной мозаики обозначают так: Первый член показывает, что этот В. содержит РНК(рибонуклеиновая кислота) (R) и она в нём однониточная (1); второй член — молекулярную массу РНК(рибонуклеиновая кислота) в миллионах и % РНК(рибонуклеиновая кислота) в частице; третий — что форма этой частицы удлинённая с параллельными сторонами и концы не закруглены, а также что подобную форму имеет и нуклеокапсид; последний член указывает, что вирус поражает высшие растения (S) и распространяется без переносчика (0).
Лит.: Рыжков В. Л., Вирусы, в сб.(сборник): Глазами ученого, М., 1963; Стэнли У., Вэленс Э., Вирусы и природа жизни, пер.(перевод) с англ.(английский), М., 1963; Вирусология и иммунология, под ред. Л. А. Зильбера, М., 1964; Молекулярные основы биологии вирусов, М., 1966; Стент Г., Молекулярная биология вирусов бактерий, пер.(перевод) с англ.(английский), М., 1965; Эндрюс К., Естественная история вирусов, пер.(перевод) с англ.(английский), М., 1969; The viruses, ed. F. М. Burnet and W. М. Stanley, v. 1—3, N. Y. — L., 1959; Fenner F. J., The biology of animal viruses, v. 1, N. Y., 1968; Gibbs A., Plant virus classification, «Advances in Virus Research», 1969, v. 14, p. 263—328.