Центральна нервова система, основна частина нервової системи тварин і людини, що складається із скупчення нервових клітин (нейронів) і їх відростків; представлена у безхребетних системою тісно зв'язаних між собою нервових вузлів (гангліїв), у хребетних тварин і людини спинним і головним мозком. Головна і специфічна функція Ц. н. с. — здійснення простих і складних високодиференційованих відбивних реакцій, що отримали назву рефлексів . У вищих тварин і людини нижчі і середні відділи Ц. н. с. — спинний мозок, довгастий мозок, середній мозок, проміжний мозок і мозочок — регулюють діяльність окремих органів і систем високорозвиненого організму, здійснюють зв'язок і взаємодію між ними, забезпечують єдність організму і цілісність його діяльності. Вищий відділ Ц. н. с. — кора великих півкуль головного мозку і найближчі підкіркові утворення — в основному регулює зв'язок і взаємини організму як єдиного цілого з довкіллям.
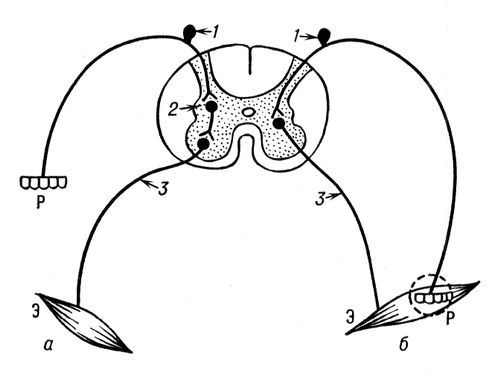
Основні межі будови і функції. Ц. н. с. пов'язана зі всіма органами і тканинами через периферичну нервову систему, яка в хребетних включає черепномозкові нерви, що відходять від головного мозку, і спинномозкові нерви — від спинного мозку, міжхребцеві нервові вузли, а також периферичний відділ вегетативної нервової системи — нервові вузли, з відповідними до них (прегангліонарними) і такими, що відходять від них (постгангліонарними) нервовими волокнами. Чутливі, або аферентні, нервові волокна, що приводять, несуть збудження в Ц. н. с. від периферичних рецепторів; по еферентних (руховим і вегетативним) нервових волокнах, що відводять, збудження з Ц. н. с. прямує до кліток виконавчих робочих апаратів (м'язи, залози судини і т.д.). У всіх відділах Ц. н. с. є аферентні нейрони, що сприймають роздратування, що приходять з периферії, і еферентні нейрони, що посилають нервові імпульси на периферію до різних виконавських еффекторним органів. Аферентні і еферентні клітки своїми відростками можуть контактувати між собою і складати двохнейронну рефлекторну дугу, що здійснює елементарні рефлекси (наприклад, сухожильні рефлекси спинного мозку). Але, як правило, в рефлекторній дузі між аферентними і еферентними нейронами розташовані вставні нервові клітини, або інтернейрони ( мал. 1 ). Зв'язок між різними відділами Ц. н. с. здійснюється також за допомогою безлічі відростків аферентних, еферентних і вставних нейронів цих відділів, створюючих внутрішньоцентральні короткі і довгі провідні дороги. До складу Ц. н. с. входять також клітки нейроглії, які виконують в ній опорну функцію, а також беруть участь в метаболізмі нервових клітин.
Рефлекторний принцип роботи Ц. н. с., як основний принцип її діяльності, до 19 ст був експериментально встановлений і вивчений лише відносно діяльності відділів, розташованих нижче за рівень великих півкуль головного мозку. Були виявлені загальні закономірності і пристосовний характер рефлекторній діяльності Ц. н. с., а також специфічні, приватні особливості функцій її різних відділів. У 19 ст І. М. Сеченовим і деякими ін. прогресивними ученими була висловлена смілива ідея про рефлекторну природу діяльності також і вищих відділів Ц. н. с., у тому числі і психічній діяльності. Ета передова, матеріалістична ідея послужила основою для класичних експериментальних і теоретичних досліджень І. П. Павлова, що привели до створення ним учення про вищій нервовій діяльності . Павлов встановив, що рефлекси, здійснювані у високорозвинених організмів корою великих півкуль головного мозку, на відміну від рефлексів всіх відділів Ц, що пролягають нижче. н. с., не природжені, а формуються в течію індивідуальному життю організму в процесі його взаємодії з довкіллям і забезпечують його найбільш досконале пристосування до умов існування. Цей новий клас рефлексів, на відміну від природжених, або безумовних рефлексів, Павлов назвав умовними рефлексами .
В протилежність первинним, примітивним уявленням про рефлекс як про трафаретну, машиноподібну, до того ж пасивну форму у відповідь діяльності Ц. н. с., класики нейрофізіології Е. Пфлюгер, Сеченов, Павлов, Ч. Шеррінгтон і ін. встановили, що і безумовні і особливо умовні рефлекси характеризуються значною динамічністю і варіабельностью: рефлекторні реакції на одні і ті ж подразники залежать від умов зовнішнього і внутрішнього середовища організму і від функціонального стану самої Ц. н. с. Основні закономірності діяльності Ц. н. с. пов'язані в першу чергу з особливостями рефлекторної дуги — структурної основи кожного рефлекторного акту. Рефлекторна дуга проводить збудження лише в одному напрямі — від рецепторного кінця до виконавського органу. Це обумовлено властивою всім нервовим клітинам структурною і функціональною поляризацією: на кінцевих розгалуженнях аксонів кожного нейрона існують мікроструктурні утворення, т.з. синапси, за допомогою яких він контактує з тілами або дендритом ін. нейронів і однобічно передає їм свою активність (Бела — Мажанді закон ) . Багатообразні зовнішні і внутрішні рецептори організму, спеціалізовані в процесі еволюції до тонкого і досконалого сприйняття окремих, якісно специфічних видів енергії, — світловою, звуковою, тепловою, механічною і хімічною, трансформують їх в процес нервового збудження, яке у вигляді ритмічних імпульсів передається послідовно від одних ланок рефлекторної дуги до інших. Збудження на своїй багатоетапній дорозі до кінцевої ланки зазнає значні зміни в ритмі, інтенсивності, швидкості і характері. У виконавських органах рефлекторне збудження може породжувати багатообразні ефекти, обумовлені специфічними особливостями структури і функцій самих виконавських органів (м'язів, залоз, судин і т.д.).
На відміну від простого нервового провідника функціональні особливості центральних відділів рефлекторної дуги характеризуються порівняно повільним виникненням і протіканням в них процесу збудження і фазових коливань рівня збудливості, що породжуються хвилями збудження. Ета своєрідна функціональна «інертність» — відносне тривале збереження стану підвищеної збудливості — обумовлює, зокрема, явище суммациі (коли слабкі неефективні стимуляції при повторенні стають ефективними), а також близьке йому явище т.з. полегшення. Центральні відділи рефлекторної дуги здатні змінювати, трансформувати ритм роздратувань. Вони відповідають на роздратування рецепторів, як правило, своїм власним, властивим ним ритмом хвиль збудження: то вищим, ніж ритм роздратування, то нижчим; при цьому між силою роздратування і інтенсивністю рефлекторної реакції є лише відносна відповідність, спостережувана зазвичай при дії подразників помірної сили. Якщо діють сильні і тривалі роздратування, нервові центри приходять в стан гальмування, в цьому випадку слабке роздратування починає викликати більший рефлекторний ефект, чим сильне (т.з. парадоксальні стосунки). Для центральних відділів рефлекторної дуги характерні також відносно висока стомлюваність і виразно виражена чутливість до біохімічних змін внутрішнього середовища організму, особливо до недоліку кисню в крові і до деяких специфічних отрут. Всі ці особливості в основному обумовлені властивостями синапсів, дендриту і тіл самих нейронів.
Для простоти рефлекторна дуга зазвичай зображається у вигляді ланцюга з ряду одиночних кліток різного роду: рецепторної клітки, аферентної, вставної і еферентної нервових клітин і виконавчої клітки. Насправді рефлекторна дуга об'єднує безліч таких ланцюгів, специфічні ланки яких є не одиночною кліткою того або іншого роду, а ансамблем взаємозв'язаних однорідних кліток. При цьому сукупність рецепторних кліток формує рецептивне поле рефлексу (рефлексогенні зони ) , об'єднання виконавчих кліток створює його робочий орган, а сукупність розташованих в Ц. н. с. нейронів утворює відповідний нервовий центр . Павлов визначив нервовий центр як сукупність нервових елементів, розташованих в різних відділах Ц. н. с., тісно зв'язаних між собою, складових єдину систему і що здійснюють регуляцію тієї або іншої функції організму. Розвиваючи і конкретизуючи цю концепцію, Е. А. Асратян запропонував розглядати центральну частину дуги безумовного рефлексу як багатоповерхову структуру, що складається з ряду гілок, кожна з яких проходіт по одному з відділів Ц. н. с., що беруть участь в даному рефлексі, і характеризується специфічністю ( мал. 2 ). Ці гілки не рівноцінні за своїм значенням у виконанні рефлексів: для одних рефлексів головними є гілки одних рівнів, для інших — гілки інших рівнів. Наприклад, в дузі серцево-судинних і дихальних рефлексів головна гілка проходіт через довгастий мозок, а в дузі харчових, захисних і статевих рефлексів — через проміжний мозок. Гілка подібної багатоповерхової дуги, що проходить через кору великого мозку, відповідає тому, що Павлов назвав кірковим представництвом безумовного рефлексу, і служить підставою для освіти відповідних умовних рефлексів.
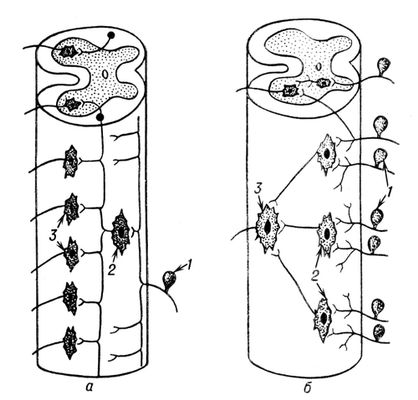
Координація рефлекторних реакцій. Ц. н. с. здійснює безліч всіляких рефлексів, що відрізняються один від одного рецепторними, центральними і еффекторнимі органами, родом, характером, знаком, походженням, мірою складності і т.п. Проте вони виникають і протікають не розрізнено і хаотично, а координований — в певному складі, порядку і послідовності відповідно до поточних потреб організму, тобто як об'єднана цілісна пристосовна діяльність. Координаційна діяльність Ц. н. с. обумовлена її структурними і функціональними особливостями. Міра генетичної спорідненості між різними структурними елементами Ц. н. с. і характер зв'язку між ними — важливі передумови їх взаємодії і координації рефлексів. Особливо істотні в цьому відношенні структурні особливості, звані розбіжністю і сходженням доріг. При розбіжності, або дивергенції, доріг кожен аферентний нейрон за допомогою безлічі кінцевих розгалужень аксонів контактує з великим числом еферентних нейронів безпосередньо або через проміжний нейрон ( мал. 3 , а). Це створює передумови для активування одним аферентним нейроном безлічі близьких до нього і віддалених еферентних нейронів і пов'язаних з ними рефлексів в певній послідовності. Наприклад, у кішки, позбавленої великих півкуль головного мозку, при слабкій стимуляції підошви задньої лапи спочатку з'являється рефлекс згинання гомілковостопного суглоба; при поступовому посиленні стимулюючого струму один за іншим до нього приєднуються рефлекси згинання колінного і тазостегнового суглобів тієї ж кінцівки, розгинальний рефлекс протилежної задньої лапи, розгинальний рефлекс однойменної передньої кінцівки і, нарешті, рефлекс згинання протилежної передньої кінцівки. Це відбувається в результаті поступового поширення (іррадіації) збудження, що виникло в аферентній ланці початкового рефлексу, на пов'язані з ним структури близьких і віддалених рефлексів спинного мозку за допомогою коротких і довгих внутріспінальних провідних доріг. При сходженні, або конвергенції, доріг декількох аферентних нейронів за допомогою кінцевих розгалужень аксонів контактують з одним еферентним нейроном безпосередньо або через проміжні нейрони ( мал. 3 , би). У останньому випадку гілки ряду аферентних нейронів сходяться до одного інтернейрону, який служить своєрідним «загальним дорогою», а гілки декілька таких інтернейронов, у свою чергу, сходяться до одного еферентного нейрона або їх ансамблю як до «загальної кінцевої дороги», яка «заблокована» для ін. потоків збудження. Конвергенція доріг в Ц. н. с. створює структурну передумову для використання одного еферентного нейрона багатьма одно- і різнорідними аферентними нейронами, число яких перевищує число еферентних у декілька разів. Наприклад, якщо роздільні роздратування слабким струмом двох різних ділянок рецептивного поля чесательного рефлексу не викликають належних рефлексів, то одночасне роздратування обох ділянок струмом тієї ж сили приводить до появи чіткого рефлексу в результаті суммациі підпорогових збуджень. Взаємне посилення рефлексів відбувається також при одночасному роздратуванні рецептивних полів двох різних рефлексів, що мають загальну кінцеву дорогу і що викликають один і той же ефект. Явища дивергенції і конвергенції спочатку були виявлені і вивчені в діяльності спинного мозку, але в подальшому було встановлено, що вони властиві і ін. відділам Ц. н. с., особливо корі великих півкуль головного мозку. У вищих відділах Ц. н. с. різко переважають аферентні нервові елементи над еферентними і тому вельми яскраво виражений принцип сходження безлічі доріг до сенсрмоторної області як до основного кортікальному «загальній дорозі». Це служить структурною передумовою для участі афферентов в здійсненні різного роду рухових безумовних рефлексів і для вироблення на їх основі багатообразних умовних рефлексів. Новітні електрофізіологічні методики дозволили багатьом сучасним нейрофізіологам (італійський фізіолог Дж. Моруцци, П. Р. Костюк і ін.) виявити і досліджувати на рівні окремих нейронів мікроструктурні і мікрофункціональні основи і механізми дивергенції і конвергенції, особливо в нейронах спинного мозку, ретикулярною формації, зорових горбів, кори головного мозку і ін. відділів Ц. н. с. Встановлено, що на телі і дендриті кожного нейрона ретикулярної формації і кори головного мозку існують синаптічеськие контакти безлічі ін. нейронів, що активуються подразниками різних модальностей і що роблять як збуджуючий, так і гальмівний вплив на «загальну дорогу».
В координації рефлексів і в діяльності Ц. н. с. в цілому важливе значення мають такі чинники, як вихідний функціональний стан залучених в даний процес нервових структур, сила, біологічне значення і тривалість дії зовнішнього стимулюючого агента, вплив внутрішніх нервових і гуморальних чинників і т.п. Проте головна функціональна основа координації рефлекторної діяльності полягає в здатності Ц. н. с. процесом збудження активувати одні синаптічеськие контакти і дороги і (що особливо важливе) одночасно блокувати процесом гальмування ін. синаптічеськие контакти і дороги в самих багатообразних комбінаціях і просторово-часових співвідношеннях, відповідно поточним потребам організму і готівковим обставинам в зовнішньому середовищі. Найважливіша роль гальмування в координаційній діяльності Ц. н. с. наочно виявляється в антагоністичних взаємодіях рефлексів, особливо у випадках, коли одночасно стимулюються різні рецептори або рецептивні поля, що мають загальний виконавський орган, але зухвалі різнорідні рефлекси. Такого типа взаємогальмуючі стосунки існують між локомоторними, захисними і познимі рефлексами, в яких беруть участь одні і ті ж розгинальні мотонейрони з іннервіруємимі ними м'язами кінцівок. Кожен з цих рефлексів може сепаратно виникати і безперешкодно протікати, якщо стимулюється лише його рецептивне поле. Але коли під час одного з рефлексів стимулюється також рецептивне поле іншого, а тим більше обидва рефлексів, то виникає конфліктна ситуація: відбувається боротьба за оволодіння загальним для них «кінцевим дорогою». Переважає той рефлекс, рецептивне поле якого в даний момент стимулюється сильнішим і який в даних умовах опиняється потрібнішим організму. Останні рефлекси блокуються гальмівним процесом і тим самим створюють сприятливі умови для безперешкодного здійснення рефлексу, що «переміг». Ін.(Древн) прикладом такої координації рефлексів може служити і т. н. реципрокная іннервація антагоністичних м'язів кінцівок, дихательних і деяких ін. систем. Як було показано російськими ученими П. А. Спіро і Н. Е. Введенським, Шеррінгтоном і ін., рефлекторне збудження мотонейронів згинальних м'язів кінцівок закономірно супроводиться гальмуванням мотонейронів їх анатомічних антагоністів — розгинальних м'язів і навпаки. Антагоністичні взаємини можуть виявлятися також рефлексами, що відносяться до різних функціональних систем; наприклад, при ковтальному акті короткочасно затримуються дихальні рухи. Найважливіша роль гальмування в координаційній діяльності Ц. н. с. наочно просліджується при введенні в організм піддослідної тварини певної дози стрихніну, тимчасово позбавляючого Ц. н. з, здібності розвивати гальмування. В такої тварини практично зникає всяка координація рефлексів: будь-яке локальне роздратування викликає одночасну рухову реакцію всієї мускулатури тіла.
Дослідженнями Сеченова і голландського фізіолога Р. Магнуса було виявлене значення вихідного функціонального стану Ц. н. с. для її координаційної діяльності. Роздратування певного рецептивного поля задньої кінцівки тварини з перерізаним спинним мозком викликає протилежні ефекти залежно від вихідного стану дратованої кінцівки: якщо кінцівка знаходиться в розігнутому стані, то роздратування викликає згинальний рефлекс, якщо ж вона зігнута, то роздратування викликає розгинальний рефлекс. Вихідне положення кінцівки через відповідні шкірні і м'язові нервові дороги відбивається в центральному органі; відбувається зміна його функціонального стану. Рецептори, закладені у виконавських органах, особливо в руховому апараті, інформують адекватні структури Ц. н. с. не лише про вихідний стан виконавського органу у спокої, але і про характер, інтенсивність, тривалість і всю динаміку виконуваної ним роботи. Безперервний потік інформації, що поступає від власних рецепторів виконавських органів в процесі їх роботи, грає важливу роль в корекції і саморегуляції роботи відповідно до поточних потребами організму і готівковою обстановкою. Цей давно відомий у фізіології Ц. н. с. важливий принцип рефлекторної саморегуляції функцій організму був в подальшому визначений як один з основних принципів кібернетики під назву зворотному зв'язку .
Своєрідна кільцева взаємодія між центральною і периферичною ланками рефлекторної дуги виявляється інколи в формі взаємного активування, створення як би кільцевого обертання збудження усередині елементів дуги, породження особливого типа ланцюга рефлексів: рефлекторне скорочення м'яза стимулює закладені в ній рецептори, це викликає нове рефлекторне скорочення того ж м'яза і т.д. (Ч. Белл, А. Ф. Самойлов ) . Подібне кільце взаємодія існує і між нейронами центральної ланки рефлекторної дуги і виявляється в декількох формах. Прикладом може служити встановлений німецьким ученим Б. Реншоу феномен. Аксон рухового нейрона спинного мозку направляє в спинний мозок поворотну гілку, яка через посредство гальмівного інтернейрона вступає в контакт з тим же руховим нейроном і робить на нього гальмівний вплив в разі надмірного інтенсивного його збудження. Подібне явище «негативному зворотному зв'язку» описане стосовно пірамідних нервових клітин кори великих півкуль головного мозку і до кліток ін. утворень Ц. н. с. Прикладом же «позитивного зворотного зв'язку» може служити описане американським ученим Р. Лоренте де Але зростаюче посилення нервового збудження завдяки тривалій його циркуляції по багатогіллястому замкнутому кільцю нейронів рефлекторного центру: поворотна гілка аксона одного з цих нейронів контактує з одним або декількома інтернейронамі, які, контактуючи з вихідним нейроном, замикають круг і створюють структурну основу для багатократної циркуляції збудження і своєрідного «самоусиленія».
взаємодії Співдружності і антагоністичних рефлексів нерозривно зв'язані між собою і по суті є різними сторонами єдиного координаційного процесу. За наявності природженої «загальної кінцевої дороги» рефлекси співдружності підсумовуються і взаємно підсилюють один одного, а антагоністичні ведуть боротьбу за цю дорогу, і рефлекс, що «переміг», на певний період часу загальмовує своїх конкурентів. В деяких випадках, наприклад під впливом зовнішнього або внутрішнього чинника (локального механічного тиску, гормону, специфічної отрути, поляризуючого струму і т.п.), значно і стійко підвищується збудливість центрального апарату якого-небудь рефлексу і цей апарат тимчасово стає своєрідним магістральним «загальним кінцевим дорогою»: стимуляція рецептивних полів різнорідних рефлексів, замість адекватних їм специфічних рефлексів, викликає або підсилює рефлекс, збудливість центру якого була підвищена заздалегідь. Ця форма координації рефлексів була виявлена і вивчена в декількох модифікаціях, відомих в сучасній нейрофізіології під назву торування доріг, суммационного рефлексу, полегшення, домінанти і т.п. Вважається, що нервове вогнище в стані підвищеної збудливості або збудження робить гальмівний вплив на певні елементи центральних структур різнорідних рефлексів, відхилює те, що йде по початкових ланках їх дуги збудження від звичайного маршруту, як би притягує до себе, підсумовує з власним збудженням і породжує або підсилює рефлекс, важливий для організму на даному етапі. Координація рефлексів залежить також від функціонального стану їх центрального апарату; наприклад, зниження його збудливості при стомленні, дії окремих нейро-гуморальних чинників і специфічних отрут веде до того, що рефлекси, що навіть домінували раніше, легко поступаються «загальною кінцевою дорогою» ін. рефлексам. Т. о., в основі багатообразних форм координації рефлексів, як і координаційній діяльності Ц. н. с. взагалі, лежать активні за своєю природою і протилежні по своїх ефектах нервові процеси — збудження і гальмування, різні комбінації і варіації розподілу їх в складних макро- і мікроструктурних елементах Ц. н. с., складна динаміка, взаємне проникнення, боротьба і взаємодія цих процесів.
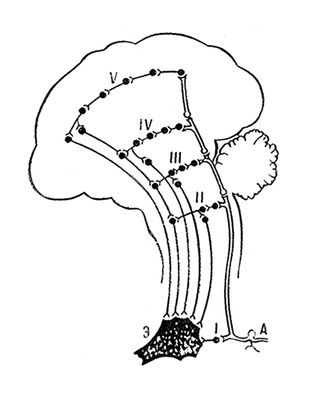
Субординація, спеціалізація і локалізація функцій. Для діяльності Ц. н. с. характерна функціональна субординація, тобто певна соподчиненность між окремими її органами, що виникла в процесі тривалої еволюції. Неоднорідність структури і нерівнозначність функцій в Ц. н. с., а також соподчиненность між її частинами намічаються на досить ранніх стадіях історичного розвитку організмів. Центральні нервові утворення і рецептори голови розвиваються швидше за центральні нервові утворення і рецептори ін. частин тіла. Розвиток провідного відділу Ц. н. с. відбувається не лише по лінії збільшення його маси, безперервній диференціації структури і спеціалізації функцій що вже сформувалися його відділів, але і по лінії розвитку в нім все нових центральних утворень з рефлекторною діяльністю більш високого рівня і важливішого значення, що очолюють і регулюючих діяльність всіх лежачих нижче за їх частини Ц. н. с. Цей процес вдосконалення, спеціалізації, локалізації і субординації функцій Ц. н. с. досягає найбільш високого рівня у вищих ссавців тварин і особливо у людиноподібних мавп і людини.
Ц. н. с. знаходиться в стані тонусу або тонічного збудження навіть за відсутності яких-небудь видимих зовнішніх ознак її активності. Тонічне збудження Ц. н. с. — прояв загальної робочої готовності і сприятливий функціональний фон для виникнення і протікання активної діяльності. У тонічному збудженні Ц. н. с. наочно виявляється субординація між її утвореннями, особливо в довгастому, середньому і проміжному мозку. Наприклад, у вищих тварин хірургічне перерізання або холодовая блокада доріг між довгастим і спинним мозком, тобто припинення субординаційного впливу першого на другій, спричиняє за собою розвиток спінального шоку — глибоке і тривале пригноблення рефлексів спинного мозку. Наслідком подібного перерізання на рівні передніх горбів четверохолмія буває т.з. децеребрационная ригідність — сильне тонічне скорочення всіх розгинальних м'язів і втрата твариною здатності активно приймати нормальну позу або підтримувати її.
Ще яскравіше субординація між відділами Ц. н. с. виявляється в процесі її активної діяльності. При цьому кожна вищерозміщена ланка Ц. н. с. здійснює складніші по структурі і складу рефлекси, досконалішу їх інтеграцію із залученням також рефлексів, регульованих ланками, що пролягають нижче. Особливості рефлекторної діяльності основних «поверхів» Ц. н. с. можна представити в наступному вигляді: рефлекси сегментів спинного мозку охоплюють лише окремі органи (наприклад, окремі кінцівки); досконаліші рефлекси довгастого мозку поширюються на діяльність окремих систем органів (травною, дихательной, серцево-судинною, рухового апарату і т.д.); рефлекси середнього мозку охоплюють всю скелетну мускулатуру тіла і забезпечують організацію таких складних рухових функцій, як стояння і ходьба; утворення проміжного мозку рефлекторно регулюють і координують діяльність внутрішніх органів всіх систем організму у всіляких поєднаннях у зв'язку із здійснюваними ними життєво важливими безумовними рефлексами — піщедобивательнимі, захисними, статевими і т.п. Великі півкулі головного мозку здатні не лише удосконалювати всі ці рефлекси і об'єднувати їх в складніші комплекси, але і створювати якісно новий вигляд рефлексів — умовні рефлекси. При цьому чим вище рівень розвитку тварини і рівень організації його Ц. н. с., тим сильніше «влада» вищих її відділів над тими, що пролягають нижче, тим значніше їх участь в регулюванні багатообразних функцій організму і управлінні ними. Посилення в процесі еволюції значення вищих відділів Ц. н. с. в багатообразній життєдіяльності організму визначається як церебралізация, енцефалізація, або кортікалізация, функцій організму (див. Кортіко-вісцелярні стосунки ) . Проте і відділи Ц, що пролягають нижче. н. с. впливають на вищерозміщених; крім того, від низу до верху поетапно передається також вся інформація від зовнішніх і внутрішніх органів. Тому субординацію в Ц. н. с. слід розглядати лише як вираження превалюючого напряму в складній і багатообразній взаємодії між нервовими утвореннями різних рівнів. Наприклад ретикулярна формація надає сильну активуючу і гальмівну дію на функціональний стан майже всіх частин Ц. н. с., у тому числі і на кору великих півкуль головного мозку, яка у свою чергу впливає на функціональний стан і діяльність ретикулярної формації і ін. глибинних утворень мозку, у тому числі і на передавальні вузли висхідних трактів, регулюючи тим самим потік отримуваною по ним інформації. Кільцева взаємодія між утвореннями Ц. н. с. і саморегуляція їх функцій підтверджують правильність положення Павлова про провідну роль кори великих півкуль головного мозку в об'єднаній і цілісній діяльності всієї Ц. н. с.
Структурні і функціональні особливості Ц. н. с. забезпечують різноманіття і досконалість її діяльності відповідно до потреб організму, виникнення у разі потреби нових форм координації. Багатий фонд «запасних можливостей» і компенсаторних пристосувань Ц. н. с. має важливе біологічне значення як в умовах нормального існування організму, так і при всякого роду пошкодженнях периферичних сприймаючих і старанних органів, нервових структур, що приводять і відводять, і різних органів самої Ц. н. с. Про патологію Ц. н. с. див.(дивися) Нервові хвороби, Психічні хвороби, Череп .
Літ.: Прохаська Р., Фізіологія, або наука про єство людському, пер.(переведення) з йому.(німецький), СП(Збори постанов) БИ. 1822; Орбелі Л. А., Лекції з фізіології нервової системи, 3 видавництва, М. — Л., 1938; Декарт Р., Ізбр. проїзв.(твір), пер.(переведення) з франц.(французький) і лат.(латинський), М., 1950; Ухтомський А. А., Собр. соч.(вигадування), т. 1, Л., 1950; Павлов І. П., Повні збори соч.(вигадування), 2 видавництва, т. 1—6, М. — Л., 1951—52; Введенський Н. Е., Повні збори соч.(вигадування), т. 4, Л., 1953; Сеченов І. М., Ізбр. проїзв.(твір), т. 1—2, М., 1952—56; Ройтбак А. І., Біоелектричні явища в корі великих півкуль, ч. 1, Тб., 1955; Магнус Р., Установка тіла, пер.(переведення) з йому.(німецький), М. — Л., 1962; Мегун Р., Безсонний мозок, пер.(переведення) з англ.(англійський), 2 видавництво, М., 1965; Берітов І. О., Загальна фізіологія м'язової і нервової системи, 3 видавництва, т. 2, М., 1966; Самойлов А. Ф., Ізбр. праці, М., 1967; Анохин П. До., Біологія і нейрофізіологія умовного рефлексу, М., 1968; Русинів Ст С., Домінанта. Електрофізіологічні дослідження, М., 1969; Асратян Е. А., Нариси по фізіології умовних рефлексів, М., 1970; Фізіологія вищої нервової діяльності, ч. 1—2, М., 1970—71; Костюк П. Р., фізіологія центральної нервової системи, До., 1977; Ліванов М. Н., Просторова організація процесів головного мозку, М., 1972; Рабіновіч М. Я., Замикательная функція мозку, М., 1975.