Аркуш (folium), орган вищих рослин, що виконує функції фотосинтезу і транспірування, а також забезпечуючий газообмін з повітряним середовищем і що бере участь в ін. найважливіших процесах життєдіяльності рослини.
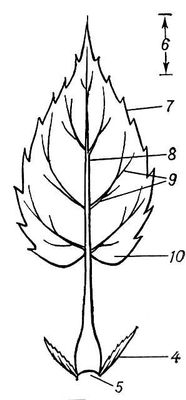
Морфологія, анатомія аркуша і його походження. Від інших вегетативних органів вищих рослин — кореня і стебла — Л. морфологічно відрізняється зазвичай дорзовентральной (а не радіальною) симетрією, бічним положенням і, як правило, обмеженим базальним зростанням. Виникає Л. із зовнішніх шарів кліток первинною мерістеми конуса наростання стебла. Л. у більшості рослин зростає спочатку верхівкою, а потім інтеркалярно. Частіше за весь Л. складається з розширеної плоскої пластинки, в якій відбуваються всі основні фізіологічні процеси, і черешка — звуженої стебльовідной частини, що прикріплює пластинку до втечі ( мал. 1 ). Завдяки черешку Л. здатний займати певне положення по відношенню до світла. В багатьох рослин (багато норічникових, губоцвітих і ін.) Л. не має вираженого черешка; такий Л. назиаєтся сидячим. Часто на одній і тій же рослині одночасно зустрічаються Л. і сидячі, і черешкові (черешкові). Нижня частина Л. незрідка розширена В т. н. піхва, що більш менш охоплює стебло. Л. з добре вираженими піхвами особливо характерні для зонтичних і злаків. Поблизу підстави Л. багатьох рослин розвиваються зазвичай парні симетрично розташовані придатки — прілістникі — у вигляді листочків, щетин, лусочок і тому подібне
Форма Л. дуже всіляка. Розрізняють Л. простій — з черешком і однією пластинкою, і Л. складний — з головним черешком, або головною віссю (рахисом), і черешками (осями) подальших порядків, до яких прикріпляються листочки; частини, що складають типовий складний Л., забезпечені зчленуваннями. Пластинка Л. формою (контуру) буває округлою, еліптичною, ланцетовидною, ромбоподібною, яйцевидною і тому подібне ( мал. 2 ). Л. може бути також голковидним, шиловидним, щетиновидним, циліндровим і іншої форми. Розрізняють, крім того, Л. лусковидною, щитовидною, воронковідной і ін. форм. Для класифікації Л. велике значення має форма підстави пластинки (серцеподібна, списоподібна, ниркоподібна і ін.), характер контура її верхівки (загострена, тупа і тому подібне) і особливості її краю (зубчастий, пильчастий, городчатий, цілісний і так далі). Залежно від характеру і глибини розчленовування пластинки розрізняють Л. лопатеві роздільні або розітнуті. Велике значення (особливо для дослідження викопних рослин ) має характер розташування жилок в пластинці, тобто жилкування (перисте, пальчате, долонеподібне, дугоподібне і ін.). Л. може бути голим, опушеним, зморшкуватим, гладким, а також плівчастим, шкірястим або соковитим. Середня довжина Л. 3—10 см, проте розміри його варіюють від декількох мм до 20 м-код (в пальми рафії смолистої). Тривалість життя Л., особливо в рослин помірних широт, зазвичай невелика — близько 6 мес; із закінченням вегетаційного періоду Л. відмирають. Лише у вічнозелених рослин Л. змінялися не періодично, а може жити більше року (див. Листопад ) . Л. однієї і тієї ж рослини зазвичай розрізняються трохи, головним чином розмірами; проте інколи спостерігається разнолістность, або гетерофіллія . Л. на стеблі розташовуються в певному порядку і розвиваються з тією або іншою послідовністю. В мн.(багато) рослин листорозміщення і орієнтування Л. сприяють максимальному використанню світла, утворюючи т.з. листову мозаїку .
Л., особливо в квіткових рослин, чуйно реагують на умови проживання і їх вагання. Вони легше за ін. органів здатні до пристосує. видозмінам, наприклад до життя в посушливих областях, а також до існування в ін. незвичайних або суворих (екстремальних) умовах (див. Метаморфоз ) . Не дивлячись на схожу у загальних рисах схему будови, різниця в особливостях Л. у представників різних відділів або класів вищих рослин значительна. Так, Л. хвойних зазвичай голчаної форми, епідерміс їх з товстою кутикулою, в м'якоті Л. проходят смоляні ходи, а провідні пучки оточені особливою, т.з. трансфузіонной, тканиною. Л. дводольних, як правило, з сітчастим жилкуванням, незрідка з розчленованою складною пластинкою. Л. однодольних найчастіше з паралельним або дугоподібним жилкуванням, незрідка лінійні і лише інколи розчленовані.
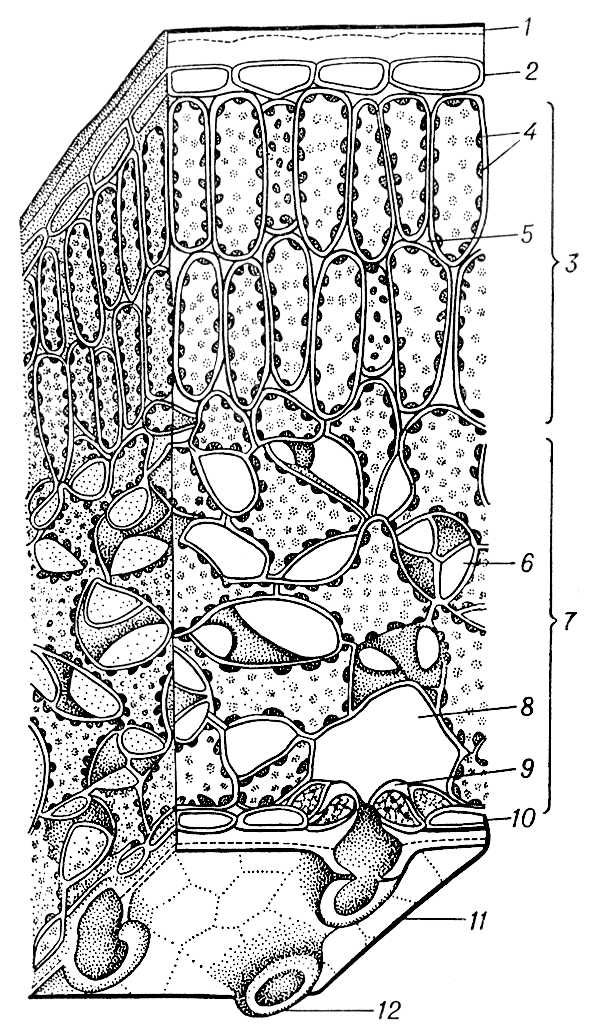
Анатомічна будова Л. розрізняється навіть в межах одного вигляду, проте у загальних рисах воно одноманітне і в різних видів ( мал. 3 ). Зазвичай пластинка Л. з обох боків забезпечена одношаровою покривною тканиною — шкіркою, або епідермісом . Між верхнім і нижнім епідермісом розташовується основна тканина, або м'якоть, Л. (мезофіл), частіше диференційована на стовпчасту, або палісадну, таку, що складається з подовжених кліток з хлоропластами і займаючу верхню, звернену до світла частину Л., і губчасту, складену з кліток з великими міжклітинниками і заповнюючу нижню частину його. Л. пронизаний жилками — провідними пучками, які поряд з механічними тканинами створюють остов Л. Механічеськие тканини, що додають Л. міцність частіше розташовуються в крупних жилок, але інколи розвиваються в м'якоті Л. Клетки епідермісу часто безбарвні, тісно примикають один до одного: вони живі і, за винятком розташованих в епідермісі устьічних кліток, позбавлені хлорофілу. Зовні епідерміс зазвичай покритий кутикулою, а інколи і восковим нальотом, що визначає захисну функцію епідермісу. Можливо, захисну роль грають і волоски на поверхні Л. Через багаточисельні устьіца в епідермісі Л. здійснюються газообмін і транспірування: у Л. поступає зовнішнє повітря, проникаюче в міжклітинників губчастої паренхіми, а з Л. виділяються пари води і гази.
Виникнення і розвиток Л. пов'язано з пристосуванням рослин до всіляких умов наземного способу життя. Найімовірніше, у більшості рослин Л. — це результат диференціювання, сплощення і зрощення бічних осей тіла рослини, що гілкується. Лише у небагатьох, наприклад в плауновідних, Л. виник як екзогенне вирощування осьових органів. Незрідка Л. вважають не окремим органом, а лише частиною єдиного цілісного органу рослин — втечі. Листоподібні вирости мохоподібних, які часто називають Л., насправді не гомологични Л. останніх вищих рослин, оскільки утворюються на гаметофіте (а не на спорофіті, як в ін. вищих рослин).
Фізіологія аркуша. Л. — активний орган, що визначає, направляє і регулюючий загальні процеси життєдіяльності рослини. Міняючи під впливом різних умов кількість і якість утворюваних в процесі фотосинтезу продуктів, Л. визначає зростання рослини і його продуктивність. Життєдіяльність Л. тісно пов'язана з життєдіяльністю рослини як цілого: забезпечуючи його асимілятами, Л. сам забезпечується за допомогою ін. органів водою, елементами мінерального живлення, деякими фізіологічно активними речовинами (наприклад, кинінамі). У відповідності з цим Л. володіє рядом важливих і характерних для виконуваних їм функцій ознак. Один з них — пристосованість до інтенсивного газообміну з повітряним середовищем. Так, 1 м 2 листової пластинки може засвоювати за годину з навколишнього повітря до 6—8 г (3—4 л ) Co 2 і одночасно виділяти в навколишнє повітря стільки ж за об'ємом O 2 . Висока ефективність забезпечується його структурою (товщина листової пластинки частіше дорівнює 0,1—0,25 мм. ) і високими показниками відношення поверхні (S) до об'єму (V) (S/v = 100—200 см 2 /см 3 ) або до сирої і сухої маси (відповідно 100 і 500 см 2 /г ) . Висока здатність Л. до газообміну визначається великою пористістю, наявністю в епідермісі устьіц (до 30 тис. на 1 см 2 ) і пористою будовою мезофіла.
Важливе умова високої фотосинтетичної активності — швидкий відтік утворюваних асимілятів в ін. органи, а у зв'язку з цим — міра розвитку провідних систем і перш за все флоеми провідних пучків (жилок). У інтенсифікації транспорту асимілятів з Л. у ін. органи грає роль в деяких рослин анатомічна структура і, зокрема, наявність обкладочних кліток провідних пучків, в яких зосереджені хлоропласти і запасаються великі кількості крохмалю. Багато рослин, Л. яких володіють такою структурою (кукурудза, цукровий очерет, сорго і ін.), характеризуються особливим типом фотосинтезу і його інтенсивністю, а також високою продуктивністю. Неодмінна умова для фотосинтезу — наявність в хлоропластах асимілюючих кліток хлорофілу і каротиноїдів, вміст яких варіює в різних рослин. При вмісті хлорофілу з розрахунку на 1 дм 2 листової пластинки 2—4 міліграма і більш кількість хлорофілу вже не лімітує фотосинтез. Л. рослин поглинають при цьому практично всю енергію вхідного в них світла з довжиною хвилі від 360 до 720 нм і деяка кількість енергії інфрачервоних променів, пропускаючи лише близько 5% енергії (12—15% зазвичай відбивається). При цьому в сонячний полудень 1 м 2 пластинки Л. може поглинути до 1,5 Мдж (360 ккал ) енергії в годині Але в кращому разі лише 5—10% цій енергії перетворюється на енергію хімічних зв'язків, остання перетворюється на тепло і, нагріваючи Л., частково вирушає в довкілля в результаті тепловіддачі. При цьому не відбувається перегріву Л., оскільки близько 247 кдж (59 ккал ) витрачається на випар кожних 100 г води, яку Л. випарює у великому кол-ве (у даному прикладі 1 м 2 листовій поверхні, утворюючи в годину при найвищій освітленості близько 4—6 г асимілятів, може випарювати до 500—700 г води). Проте часто, наприклад в зонах недостатнього водопостачання, в рослин формуються Л., будову яких забезпечує краще постачання водою, а інколи навіть більший її випар, що збільшує так звану силу, що смокче. При тимчасових порушеннях водного балансу в Л. активізуються регуляторні системи — рухи устьіц, скорочення водоотдачи клітками і так далі, що одночасно веде до ослабіння інтенсивності газообміну, а отже, і фотосинтезу. Збереженню водно-теплового балансу на оптимальному рівні сприяють також восковий наліт на Л., опушенность епідермісу, скорочення числа устьіц, потовщення Л. в результаті сильного розвитку добре оводненной тканини, перетворення Л. у колючки, луски і тому подібне (функцію фотосинтезу виконують стеблові органи, що при цьому містять хлорофіл). Зредуковані Л. і в багатьох рослин-паразитів, які харчуються органічними речовинами не за рахунок фотосинтезу, а добуваючи їх через присоски у рослин-господарів.
Проте не окремий Л. і не одиночна рослина, а їх сукупність в природних або культурних фітоценозах як цілісних фотосинтезуючих системах забезпечує найкраще використання Л. в процесі фотосинтезу потоків тієї, що приходить на поверхня Землі енергії сонячного світла, Co 2 з повітря і чинників грунтової родючості, а отже, найвищу продуктивність. За сприятливих умов фітоценози більшості рослин практично можуть поглинати всю енергію вхідного в них світла і давати максимальну продукцію, утворюючи оптимальну площу Л. — 4—5 м 2 на 1 м 2 займаної ними площі землі. Т. о., здобуття посівів і насаджень з оптимальною площею Л. і динамікою її формування — найважливіше завдання підвищення продуктивності рослин. Це завдання вирішується по-різному, але на основі загальних закономірностей фотосинтетичної діяльності Л. рослини у фітоценозах. Т. о., через фотосинтез, здійснюваний переважно в Л., реалізується космічна функція зелених рослин, які поглинають сонячну енергію, використовуючи її на біосинтез органічної речовини. Через процеси фотосинтезу і дихання Л. підтримують баланс O 2 і Co 2 в повітрі, через транспірування Л. беруть участь в круговороті води на Землі.
Використання листя в господарській діяльності людини всіляко. Л. багатьох рослин споживають (салат, щавель, кріп, капуста, лук, петрушка і так далі), з Л. блекоти, сенни, наперстянки і багатьох ін. отримують лікарські речовини. Л. ін. рослин використовують на корм з.-х.(сільськогосподарський) твариною у вигляді сіна, сінажу, силосу і свіжою трави, Л. шовковиці — на корм для тутового шовкопряда. З Л. герані, м'яти і ін. рослин отримують ефірні масла, використовувані в парфюмерній і кондитерській промисловості. З Л. деяких рослин добувають дубильні речовини. Л. чайного куща і тютюну служать сировиною в чайній і тютюновій промисловості. У тропіках Л. багатьох рослин використовують для здобуття грубого волокна, а Л. пальм — як покрівельний матеріал.
Літ.: Тімірязев До. А., Життя рослини (лекція 5), Ізбр. соч.(вигадування), т. 1, М., 1957; Серебряков І. Р., Морфологія вегетативних органів вищих рослин, М., 1952; фотосинтезуючі системи високої продуктивності, Сб., М., 1966; Нічипоровіч А. А., фотосинтез і теорія здобуття високих урожаїв, М., 1956; Федоров А. А., Цегельників М. Е., Артюшенко З. Т., Атлас по описовій морфології вищих рослин. Аркуш, М. — Л., 1956; Тахтаджян А. Л., Основи еволюційної морфології покритосеменних, М. — Л., 1964; Ботаніка, 7 видавництво, т. 1, М., 1966; Есау До., Анатомія рослин, М., 1969; фотосинтез і використання сонячної енергії, Сб., Л., 1971.
М. Е. Цегельників, А. А. Нічипоровіч, А. А. Федоров.