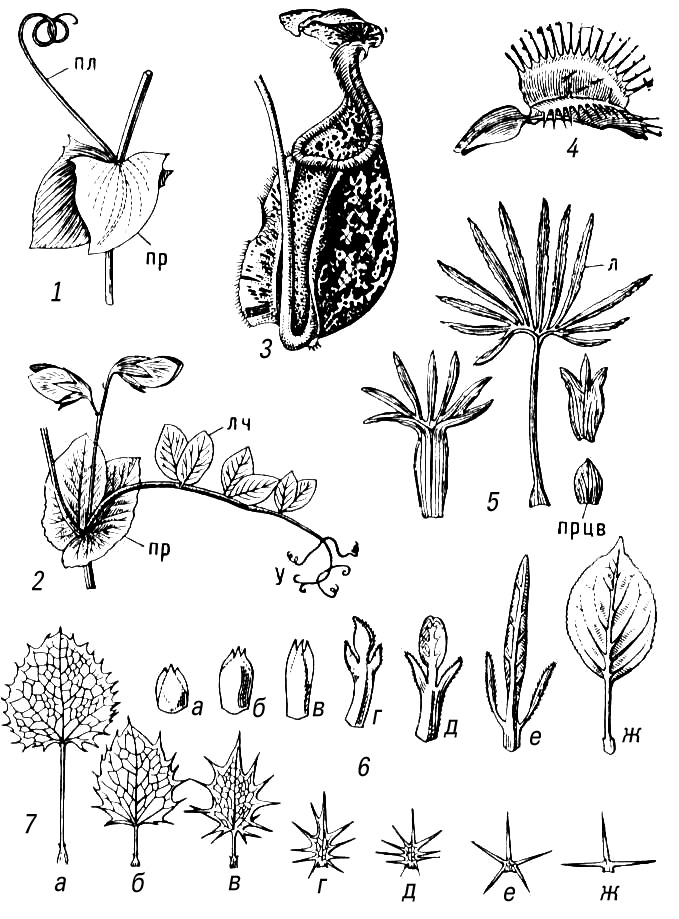
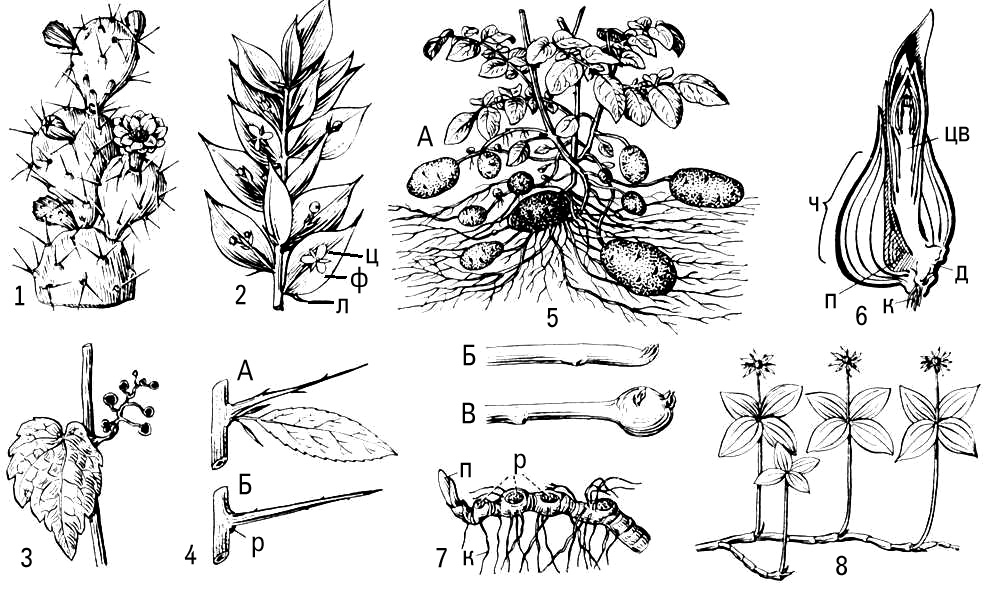
Метаморфоз (від греч.(грецький) metamórphosis — перетворення) в рослин, видозміни основних органів рослини, пов'язані зазвичай із зміною виконуваних ними функцій або умов функціонування. М. відбувається в онтогенезі рослини і полягає в зміні ходу індивідуального розвитку органу, який виробився і закріпився в процесі еволюції. М. понад усе схильні втеча в цілому і аркуш як його бічний орган, що пов'язане з різноманітністю умов середовища, що впливають на них ( мал. 1 , 2 ). Частіше за М. типової надземної втечі із зеленим листям викликаний недоліком вологи і спостерігається в рослин посушливих областей і місцепроживань. Так, в стеблових сукулентів (наприклад, кактусів і африканських молочаїв) м'ясисте стебло стало таким, що водозапасає і фотосинтезуючим органом, в пазухах недорозвиненого листя на нім розвиваються укорочені втечі з пучком колючок; завдяки безлістності в кактусів різко зменшується загальна випарювальна поверхня втечі. Зменшення випарювальної поверхні спостерігається і при таких М. надземних втеч, як кладодії (наприклад, в спаржі) і філокладії (наприклад, в рускусу). Функцію фотосинтезу в цьому випадку виконує жорстке сухувате стебло, яке незрідка стає плоским і навіть листоподібним. Інколи відбувається М. не всіх, а лише частини втеч, наприклад в дерев'янисті безлисті колючки (глід, гледічия). В ліан, що мешкають в умовах підвищеної вологості і недоліку світла, надземні втечі можуть перетворюватися у вусики — органи лазіння (наприклад, в пасифлори, винограду, в яких на вусики перетворена частина соцветій). Незрідка М. піддається лише листя (наприклад, колючки, що сидять на звичайних стеблах барбарису, вусики бобів). У вусик перетворюється або вся листова пластинка (в деяких видів чини), або лише частина листочків складного аркуша (в гороху і ін.). В комахоїдних рослин листя перетвориться в своєрідні пастки для комах. В т.з. філлодійних акацій листові пластинки можуть не розвиватися і функцію фотосинтезу виконують жорсткі черешки сплощень листя — філодії .
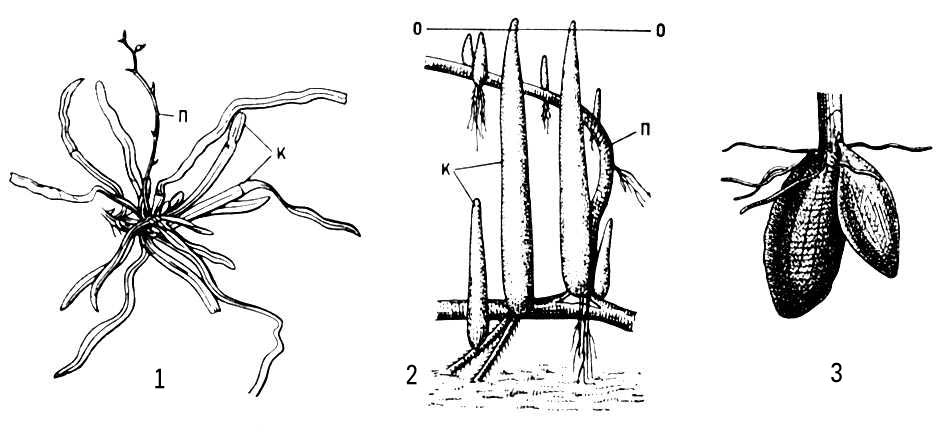
Для багатолітніх, головним чином трав'янистих, рослин звичайний М. підземних втеч, що забезпечує переживання несприятливого періоду, відновлення зростання і вегетативне розмноження. Це — запасаючі органи, що не мають зеленого листя, але забезпечені нирками: кореневища, бульби, цибулини або клубнелуковіци. М. коріння зазвичай пов'язаний з гіпертрофією запасаючої функції (наприклад, утворення коренеплодів) або із специфічною діяльністю коріння в надземному середовищі (наприклад, повітряне коріння епіфітов, дихальне коріння мангрових) ( мал. 3 ).
втечею, що Метаморфізує, пристосованою до насінного розмноження, є також квітка: чашолистки, пелюстки, тичинки і плодолистки за способом виникнення відповідають листю, а квітколоже — стеблу. Це підтверджується випадками проростання квітки (проліфікації ) , наприклад в троянди, гравілату.
Представлення о М. органів рослини складалися головним чином у зв'язку з прагненням зрозуміти природу квітки. Спроби такого роду робилися італ.(італійський) ботаніком А. Чезальпіно (16 ст), йому.(німецький) ботаніком І. Юнгом (17 ст). Термін «М-коду.» введений в науку До. Ліннєєм (1755), який помилково вважав, що частини квітки утворюються унаслідок М. різних тканин стебла. До. Ф. Вольф (1759) вперше описав формування зачатків листя і частин квітки на конусі наростання втечі і т.ч. показав їх гомологію. Учення о М. було сформульовано І. Ст Гете (1790), який розумів під М. процес зміни аркуша в ході онтогенезу рослини. Ідеї Гете були використані для пояснення утворення органів, що метаморфізували, в філогенезі різних систематичних груп рослин.
М. може відбуватися на різних етапах розвитку органу. В багатьох трав'янистих рослин втеча спочатку розташовується на поверхні землі і несе зелене асимілююче листя, а потім втрачає їх, утворює додаткове коріння і поступово занурюється в грунт, перетворюючись на запасаючий підземний орган — кореневище. Так відбувається дійсний М. — перетворення одного органу в іншій із зміною форми і функції. У більшості ж випадків метаморфізують не дорослі органи, а їх зачатки. Детерміація зачатка органу що визначає його остаточну подобу і що відбувається на різних етапах його розвитку, згідно з представленнями сов.(радянський) фізіолога Д. А. Сабініна, пов'язана з накопиченням визначених фізіологічно активних речовин і залежить від ряду зовнішніх і внутрішніх чинників.
Літ.: Серебряков І. Р., Морфологія вегетативних органів вищих рослин, М., 1952; Федоров А. А., Цегельників М. Е., Артюшенко З. Т., Атлас по описовій морфології вищих рослин, т. 1—2, М. — Л., 1956—62; Гете І. Ст, Вибрані вигадування по природознавству, М., 1957; Сабінін Д. А., Фізіологія розвитку рослин, М., 1963; Первухина Н. Ст, Проблеми морфології і біології квітки, Л., 1970.
Т. І. Серебрякова.
У тварин метаморфозом, або метаболією, називається глибоке перетворення будови організму в період постембріонального розвитку. М. пов'язаний зазвичай з різкою зміною умов існування і способу життя тварини протягом його індивідуального розвитку — онтогенезу, наприклад з переходом від свободноплавающего до прикріпленого способу життя, від водного — до наземного або повітряному і т.п. Тому в життєвому циклі тварин, що розвиваються з М., буває хоч би одна личинкова стадія, в якій організм істотно відрізняється від дорослої тварини. При розвитку с М. тварини на тих або ін. стадіях онтогенезу виконують різні функції, сприяючі збереженню і процвітанню вигляду ( мал. 4 ).
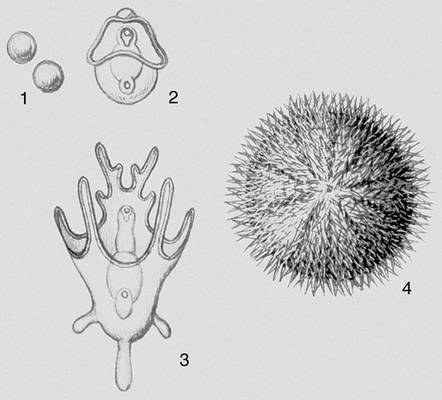
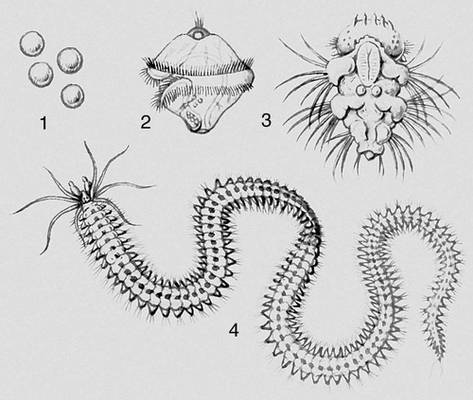
Вже в простих, наприклад в інфузорій, що смокчуть, є елементи М.: нові особини, що відгалужуються, мають війковий покрив і плавають, потім втрачають вії, стають прікрепленножівущимі і харчуються за допомогою трубочок, що витягуються. Для нижчих безхребетних (губки, кишечнополостниє) характерний М., при якому свободноплавающие личинки (паренхимула, амфібластула, планула) виконують функцію розселення вигляду. У багатьох випадках такий М. ускладнюється зміною поколінь (фаз розвитку), що розмножуються безстатевим або статевим дорогою (наприклад, в сцифомедуз, багатьох плоских черв'яків). Своєрідний т.з. некротичний М. в немертін, в яких усередині личинки розвивається майбутня доросла особина, а основна маса тіла личинки відмирає. При М. без чергування поколінь (у багатьох безхребетних) з яйця виходить личинка, що виконує расселітельную функцію (наприклад, трохофора морських многощетінкових черв'яків, велігер морських молюсків). При цьому у дорослої тварини розрізняють ларвальниє сегменти (що збереглися від першої личинки) і постларвальниє (що з'явилися пізніше); наприклад, в ракоподібних антеннули, антени і мандібули розвиваються з придатків наупліуса і відповідають ларвальним сегментам.
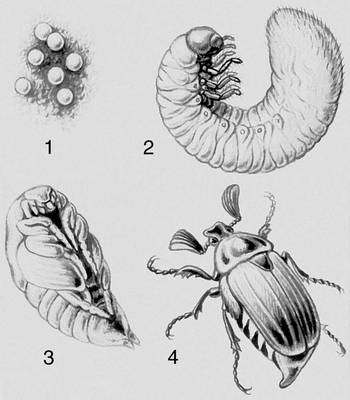
Перехід до життя в прісній воді і на суші привів до втрати личинкових стадій розвитку. Випадки, як, наприклад, у виноградного равлика, коли з яйця вилуплюється равлик, схожий на дорослу, але в яйці вона проходіт стадію, велігер морських форм, що нагадує, називається кріптометаболієй. У багатьох багатоніжок і нижчих бессяжкових комах в постембріональному періоді розвитку зміни пов'язані лише з збільшенням числа сегментів і члеників вусиків — анаморфоз. Для більшості первічнобеськрилих комах характерний розвиток без істотних змін — протометаболія. Розвиток крил у комах привів до різних змін в їх онтогенезі. Якщо спосіб життя ранніх постембріональних стадій і дорослої форми схожий, з яйця виходить личинка (німфа), схожа на доросла комаха, і зміни організації супроводяться в основному поступовим зростанням зачатків крил (геміметаболія, епіморфоз). Якщо в онтогенезі відбувається різке розділення основних функцій (живлення у стадії личинки, розселення і розмноження в дорослій стадії), то говорять про складний М. (голометаболія ) . В цьому випадку червоподібна личинка зазвичай не схожа на доросла комаха. Перехід личинки в дорослу форму супроводиться різкими змінами організму і здійснюється на стадії тієї, що не харчується, зазвичай малорухливою лялечки, в телі якої відбувається руйнування личинкових тканин і формування органів дорослої комахи (крил і ін.). Личинки голкошкірих — діплеурула, біпіннарія, плутеус і ін., а також кишководишних — торнарія, хвостата личинка асцидій — вільно плавають, виконуючи функцію розселення вигляду.
Серед хребетних М. відомий у міног, личинка яких — песькоройка — живе в грунті, а дорослі міноги — напівпаразити риб. В ряду риб, наприклад в Двоякодихаючих, личинка із зовнішніми зябрами, а у дорослих особин зябра розташовані в спеціальній порожнині, є у них також легке. В земноводних з яйця виходить личинка — пуголовок, схожа на рибку і така, що мешкає у воді. У міру М. личинкові органи втрачаються і з'являються органи дорослої тварини. Жабеня із залишком хвоста виходить на сушу і незабаром набуває подоби дорослої жаби. Регуляція М. здійснюється гормонами. У комах в 1954 виділений і в 1966 синтезований гормон проторакальних залоз — екдізон, регулюючий М. і ліньки. Затримку М. викликає ювенільний гормон прилеглих тіл. В земноводних М. регулюється гормонами щитовидної залози.
Літ.: Їжачків І. І., Метаморфоз комах, М., 1929; Гиляров М. С., Вплив характеру розселення на хід онтогенезу комах, «Журнал загальної біології», 1945, т. 6 № 1; Іванов П. П., Керівництво по загальній і порівняльній ембріології, Л., 1945; Novàk V. J. A., Insect hormones, 3 ed., L., 1966.