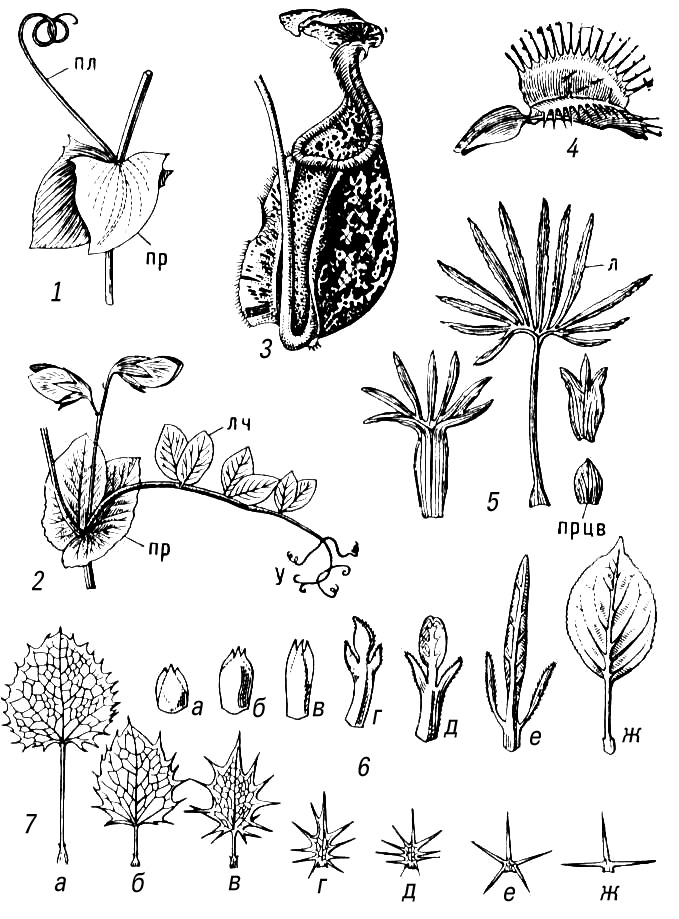
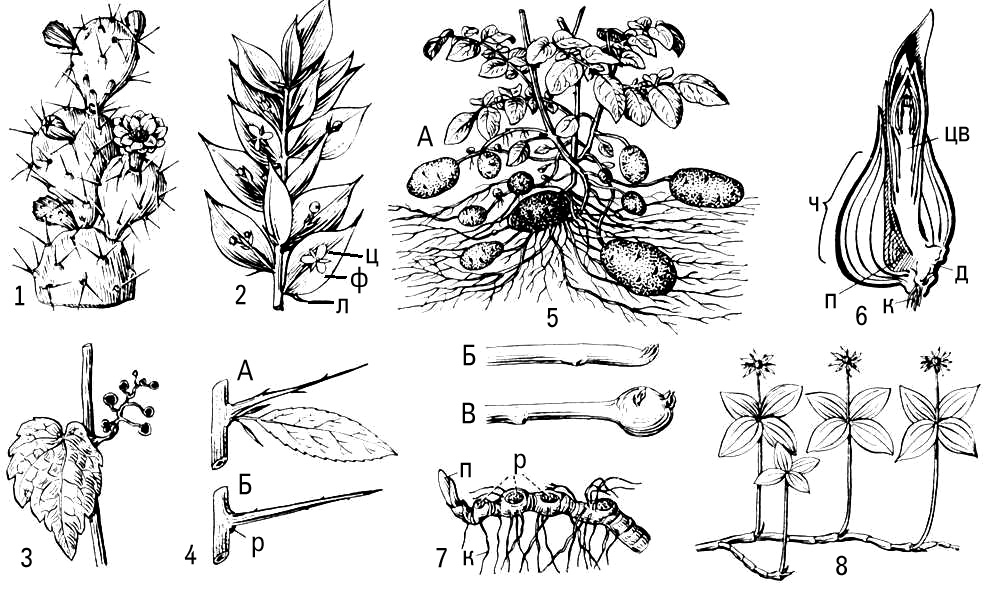
Метаморфоз (от греч.(греческий) metamórphosis — превращение) у растений, видоизменения основных органов растения, связанные обычно со сменой выполняемых ими функций или условий функционирования. М. происходит в онтогенезерастения и заключается в изменении хода индивидуального развития органа, которое выработалось и закрепилось в процессе эволюции. М. более всего подвержены побег в целом и лист как его боковой орган, что связано с разнообразием влияющих на них условий среды (рис. 1, 2). Чаще М. типичного надземного побега с зелёными листьями вызван недостатком влаги и наблюдается у растений засушливых областей и местообитаний. Так, у стеблевых суккулентов(например, кактусов и африканских молочаев) мясистый стебель стал водозапасающим и фотосинтезирующим органом, в пазухах недоразвитых листьев на нём развиваются укороченные побеги с пучком колючек; благодаря безлистности у кактусов резко уменьшается общая испаряющая поверхность побега. Уменьшение испаряющей поверхности наблюдается и при таких М. надземных побегов, как кладодии(например, у спаржи) и филлокладии (например, у иглицы). Функцию фотосинтеза в этом случае выполняет жёсткий суховатый стебель, который нередко становится плоским и даже листовидным. Иногда происходит М. не всех, а только части побегов, например в деревянистые безлистные колючки (боярышник, гледичия). У лиан, обитающих в условиях повышенной влажности и недостатка света, надземные побеги могут преобразовываться в усики — органы лазания (например, у пассифлоры, винограда, у которых в усики превращена часть соцветий). Нередко М. подвергаются только листья (например, колючки, сидящие на обычных стеблях барбариса, усики бобовых). В усик превращается или вся листовая пластинка (у некоторых видов чины), или только часть листочков сложного листа (у гороха и др.). У насекомоядных растенийлистья преобразуются в своеобразные ловушки для насекомых. У т. н. филлодийных акаций листовые пластинки могут не развиваться и функцию фотосинтеза выполняют жёсткие уплощённые черешки листьев — филлодии.
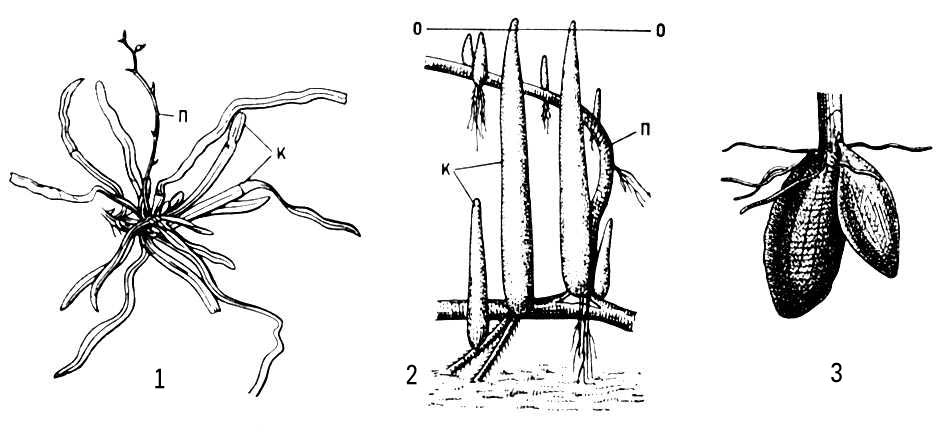
Для многолетних, главным образом травянистых, растений обычен М. подземных побегов, обеспечивающий переживание неблагоприятного периода, возобновление роста и вегетативное размножение. Это — запасающие органы, не имеющие зелёных листьев, но снабженные почками: корневища, клубни, луковицы или клубнелуковицы. М. корней обычно связан с гипертрофией запасающей функции (например, образование корнеплодов) или со специфической деятельностью корней в надземной среде (например, воздушные корни эпифитов, дыхательные корни мангровых) (рис. 3).
Метаморфизированным побегом, приспособленным к семенному размножению, является также цветок: чашелистики, лепестки, тычинки и плодолистики по способу возникновения соответствуют листьям, а цветоложе — стеблю. Это подтверждается случаями прорастания цветка (пролификации), например у розы, гравилата.
Представления о М. органов растения складывались главным образом в связи со стремлением понять природу цветка. Попытки такого рода предпринимались итал.(итальянский) ботаником А. Чезальпино (16 в.), нем.(немецкий) ботаником И. Юнгом (17 в.). Термин «М.» введён в науку К. Линнеем (1755), который ошибочно считал, что части цветка образуются вследствие М. разных тканей стебля. К. Ф. Вольф (1759) впервые описал формирование зачатков листьев и частей цветка на конусе нарастания побега и т. о. показал их гомологию. Учение о М. было сформулировано И. В. Гёте (1790), который понимал под М. процесс изменения листа в ходе онтогенеза растения. Идеи Гёте были использованы для объяснения образования метаморфизированных органов в филогенезе разных систематических групп растений.
М. может происходить на разных этапах развития органа. У многих травянистых растений побег сначала располагается на поверхности земли и несёт зелёные ассимилирующие листья, а затем теряет их, образует придаточные корни и постепенно погружается в почву, превращаясь в запасающий подземный орган — корневище. Так происходит истинный М. — превращение одного органа в другой со сменой формы и функции. В большинстве же случаев метаморфизируются не взрослые органы, а их зачатки. Детерминация зачатка органа, определяющая его окончательный облик и происходящая на разных этапах его развития, согласно представлениям сов.(советский) физиолога Д. А. Сабинина, связана с накоплением определённых физиологически активных веществ и зависит от ряда внешних и внутренних факторов.
Лит.: Серебряков И. Г., Морфология вегетативных органов высших растений, М., 1952; Федоров А. А., Кирпичников М. Э., Артюшенко З. Т., Атлас по описательной морфологии высших растений, т. 1—2, М. — Л., 1956—62; Гёте И. В., Избранные сочинения по естествознанию, М., 1957; Сабинин Д. А., Физиология развития растений, М., 1963; Первухина Н. В., Проблемы морфологии и биологии цветка, Л., 1970.
Т. И. Серебрякова.
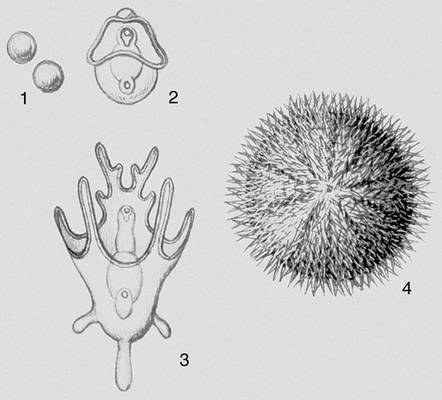
У животных метаморфозом, или метаболией, называется глубокое преобразование строения организма в период постэмбрионального развития. М. связан обычно с резкой сменой условий существования и образа жизни животного в течение его индивидуального развития — онтогенеза, например с переходом от свободноплавающего к прикрепленному образу жизни, от водного — к наземному или воздушному и т.п. Поэтому в жизненном цикле животных, развивающихся с М., бывает хотя бы одна личиночная стадия, в которой организм существенно отличается от взрослого животного. При развитии с М. животные на тех или др. стадиях онтогенеза выполняют разные функции, способствующие сохранению и процветанию вида (рис. 4).
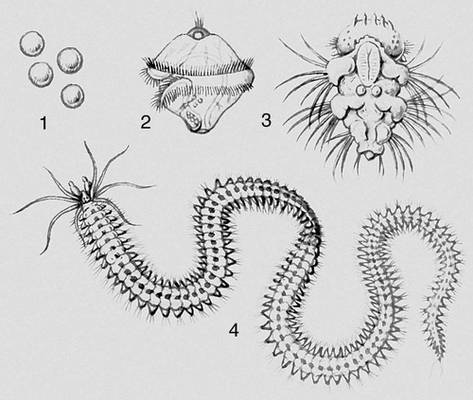
Уже у простейших, например у сосущих инфузорий, есть элементы М.: отпочковывающиеся новые особи имеют ресничный покров и плавают, затем теряют реснички, становятся прикреплённоживущими и питаются с помощью вытягивающихся трубочек. Для низших беспозвоночных (губки, кишечнополостные) характерен М., при котором свободноплавающие личинки (паренхимула, амфибластула, планула) выполняют функцию расселения вида. Во многих случаях такой М. осложняется сменой поколений (фаз развития), размножающихся бесполым или половым путём (например, у сцифомедуз, многих плоских червей). Своеобразен т. н. некротический М. у немертин, у которых внутри личинки развивается будущая взрослая особь, а основная масса тела личинки отмирает. При М. без чередования поколений (у многих беспозвоночных) из яйца выходит личинка, выполняющая расселительную функцию (например, трохофора морских многощетинковых червей, велигер морских моллюсков). При этом у взрослого животного различают ларвальные сегменты (сохранившиеся от первой личинки) и постларвальные (появившиеся позже); например, у ракообразных антеннулы, антенны и мандибулы развиваются из придатков науплиуса и соответствуют ларвальным сегментам.
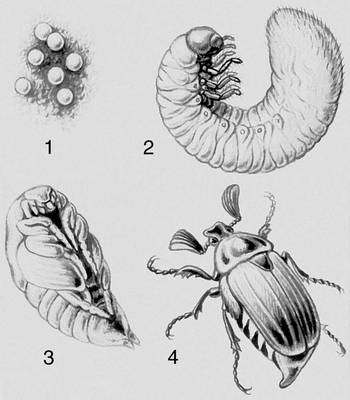
Переход к жизни в пресной воде и на суше привёл к утрате личиночных стадий развития. Случаи, как, например, у виноградной улитки, когда из яйца вылупляется улитка, похожая на взрослую, но в яйце она проходит стадию, напоминающую велигер морских форм, называется криптометаболией. У многих многоножек и низших бессяжковых насекомых в постэмбриональном периоде развития изменения связаны лишь с увеличением числа сегментов и члеников усиков — анаморфоз. Для большинства первичнобескрылых насекомых характерно развитие без существенных изменений — протометаболия. Развитие крыльев у насекомых привело к разным изменениям в их онтогенезе. Если образ жизни ранних постэмбриональных стадий и взрослой формы сходен, из яйца выходит личинка (нимфа), похожая на взрослое насекомое, и изменения организации сопровождаются в основном постепенным ростом зачатков крыльев (гемиметаболия, эпиморфоз). Если в онтогенезе происходит резкое разделение основных функций (питание в стадии личинки, расселение и размножение во взрослой стадии), то говорят о сложном М. (голометаболия). В этом случае червеобразная личинка обычно не похожа на взрослое насекомое. Переход личинки во взрослую форму сопровождается резкими изменениями организма и осуществляется на стадии непитающейся, обычно малоподвижной куколки, в теле которой происходит разрушение личиночных тканей и формирование органов взрослого насекомого (крыльев и др.). Личинки иглокожих — диплеурула, бипиннария, плутеус и др., а также кишечнодышащих — торнария, хвостатая личинка асцидий — свободно плавают, выполняя функцию расселения вида.
Среди позвоночных М. известен у миног, личинка которых — пескоройка — живёт в грунте, а взрослые миноги — полупаразиты рыб. У ряда рыб, например у двоякодышащих, личинка с наружными жабрами, а у взрослых особей жабры расположены в специальной полости, имеется у них также лёгкое. У земноводных из яйца выходит личинка — головастик, похожая на рыбку и обитающая в воде. По мере М. личиночные органы утрачиваются и появляются органы взрослого животного. Лягушонок с остатком хвоста выходит на сушу и вскоре приобретает облик взрослой лягушки. Регуляция М. осуществляется гормонами. У насекомых в 1954 выделен и в 1966 синтезирован гормон проторакальных желёз — экдизон, регулирующий М. и линьки. Задержку М. вызывает ювенильный гормон прилежащих тел. У земноводных М. регулируется гормонами щитовидной железы.
Лит.: Ежиков И. И., Метаморфоз насекомых, М., 1929; Гиляров М. С., Влияние характера расселения на ход онтогенеза насекомых, «Журнал общей биологии», 1945, т. 6, № 1; Иванов П. П., Руководство по общей и сравнительной эмбриологии, Л., 1945; Novàk V. J. A., Insect hormones, 3 ed., L., 1966.