Лист (folium), орган высших растений, выполняющий функции фотосинтеза и транспирации, а также обеспечивающий газообмен с воздушной средой и участвующий в др. важнейших процессах жизнедеятельности растения.
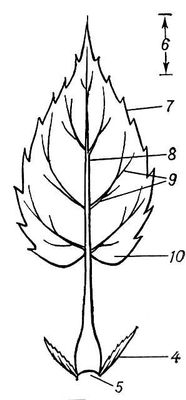
Морфология, анатомия листа и его происхождение. От других вегетативных органов высших растений — корня и стебля — Л. морфологически отличается обычно дорзовентральной (а не радиальной) симметрией, боковым положением и, как правило, ограниченным базальным ростом. Возникает Л. из наружных слоев клеток первичной меристемы конуса нарастания стебля. Л. у большинства растений растет сначала верхушкой, а затем интеркалярно. Чаще всего Л. состоит из расширенной плоской пластинки, в которой происходят все основные физиологические процессы, и черешка — суженной стеблевидной части, прикрепляющей пластинку к побегу (рис. 1). Благодаря черешку Л. способен занимать определённое положение по отношению к свету. У многих растений (многих норичниковых, губоцветных и др.) Л. не имеет выраженного черешка; такой Л. назыается сидячим. Часто на одном и том же растении одновременно встречаются Л. и сидячие, и черешчатые (черешковые). Нижняя часть Л. нередко расширена в т. н. влагалище, более или менее охватывающее стебель. Л. с хорошо выраженными влагалищами особенно характерны для зонтичных и злаков. Близ основания Л. многих растений развиваются обычно парные симметрично расположенные придатки — прилистники — в виде листочков, щетинок, чешуек и т.п.
Форма Л. очень разнообразна. Различают Л. простой — с черешком и одной пластинкой, и Л. сложный — с главным черешком, или главной осью (рахисом), и черешками (осями) последующих порядков, к которым прикрепляются листочки; части, слагающие типичный сложный Л., снабжены сочленениями. Пластинка Л. по форме (очертанию) бывает округлой, эллиптической, ланцетовидной, ромбовидной, яйцевидной и т.п. (рис. 2). Л. может быть также игловидным, шиловидным, щетиновидным, цилиндрическим и иной формы. Различают, кроме того, Л. чешуевидной, щитовидной, воронковидной и др. форм. Для классификации Л. большое значение имеет форма основания пластинки (сердцевидная, копьевидная, почковидная и др.), характер очертания её верхушки (заострённая, тупая и т.п.) и особенности её края (зубчатый, пильчатый, городчатый, цельный и т.д.). В зависимости от характера и глубины расчленения пластинки различают Л. лопастные, раздельные или рассеченные. Большое значение (особенно для исследования ископаемых растений) имеет характер расположения жилок в пластинке, т. е. жилкование (перистое, пальчатое, дланевидное, дуговидное и др.). Л. может быть голым, опушенным, морщинистым, гладким, а также плёнчатым, кожистым или сочным. Средняя длина Л. 3—10 см, однако размеры его варьируют от нескольких мм до 20 м (у пальмы рафии смолистой). Продолжительность жизни Л., особенно у растений умеренных широт, обычно невелика — около 6 мес; с окончанием вегетационного периода Л. отмирают. Лишь у вечнозелёных растений Л. сменяются не периодически, а могут жить более года (см. Листопад). Л. одного и того же растения обычно различаются незначительно, главным образом размерами; однако иногда наблюдается разнолистность, или гетерофиллия. Л. на стебле располагаются в определённом порядке и развиваются с той или иной последовательностью. У мн.(многие) растений листорасположениеи ориентировка Л. способствуют максимальному использованию света, образуя т. н. листовую мозаику.
Л., особенно у цветковых растений, чутко реагируют на условия обитания и их колебания. Они легче др. органов способны к приспособит. видоизменениям, например к жизни в засушливых областях, а также к существованию в др. необычных или суровых (экстремальных) условиях (см. Метаморфоз). Несмотря на сходную в общих чертах схему строения, разница в особенностях Л. у представителей разных отделов или классов высших растений значительна. Так, Л. хвойных обычно игольчатой формы, эпидермис их с толстой кутикулой, в мякоти Л. проходят смоляные ходы, а проводящие пучки окружены особой, т. н. трансфузионной, тканью. Л. двудольных, как правило, с сетчатым жилкованием, нередко с расчленённой сложной пластинкой. Л. однодольных чаще всего с параллельным или дуговидным жилкованием, нередко линейные и лишь иногда расчленённые.
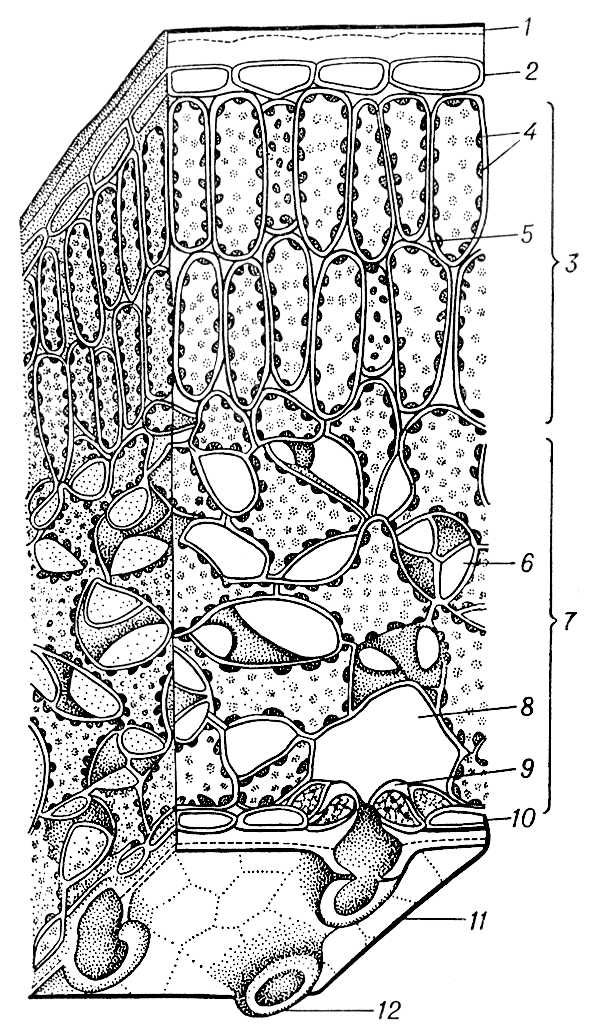
Анатомическое строение Л. различается даже в пределах одного вида, однако в общих чертах оно единообразно и у разных видов (рис. 3). Обычно пластинка Л. с обеих сторон снабжена однослойной покровной тканью — кожицей, или эпидермисом. Между верхним и нижним эпидермисом располагается основная ткань, или мякоть, Л. (мезофилл), чаще дифференцированная на столбчатую, или палисадную, состоящую из удлинённых клеток с хлоропластами и занимающую верхнюю, обращенную к свету часть Л., и губчатую, сложенную из клеток с большими межклетниками и заполняющую нижнюю часть его. Л. пронизан жилками — проводящими пучками, которые наряду с механическими тканями создают остов Л. Механические ткани, придающие Л. прочность, чаще располагаются у крупных жилок, но иногда развиваются в мякоти Л. Клетки эпидермиса часто бесцветные, тесно примыкают друг к другу: они живые и, за исключением расположенных в эпидермисе устьичных клеток, лишены хлорофилла. Снаружи эпидермис обычно покрыт кутикулой, а иногда и восковым налётом, что определяет защитную функцию эпидермиса. Возможно, защитную роль играют и волоски на поверхности Л. Через многочисленные устьица в эпидермисе Л. осуществляются газообмен и транспирация: в Л. поступает наружный воздух, проникающий в межклетники губчатой паренхимы, а из Л. выделяются пары воды и газы.
Возникновение и развитие Л. связано с приспособлением растений к разнообразным условиям наземного образа жизни. Вероятнее всего, у большинства растений Л. — это результат дифференцировки, уплощения и срастания боковых осей ветвящегося тела растения. Лишь у немногих, например у плауновидных, Л. возник как экзогенный вырост осевых органов. Нередко Л. считают не отдельным органом, а лишь частью единого цельного органа растений — побега. Листовидные выросты мохообразных, которые часто называют Л., на самом деле не гомологичны Л. остальных высших растений, т.к. образуются на гаметофите (а не на спорофите, как у др. высших растений).
Физиология листа. Л. — активный орган, определяющий, направляющий и регулирующий общие процессы жизнедеятельности растения. Меняя под влиянием различных условий количество и качество образуемых в процессе фотосинтеза продуктов, Л. определяет рост растения и его продуктивность. Жизнедеятельность Л. тесно связана с жизнедеятельностью растения как целого: снабжая его ассимилятами, Л. сам снабжается при помощи др. органов водой, элементами минерального питания, некоторыми физиологически активными веществами (например, кининами). В соответствии с этим Л. обладает рядом важных и характерных для выполняемых им функций признаков. Один из них — приспособленность к интенсивному газообмену с воздушной средой. Так, 1 м2 листовой пластинки может усваивать за час из окружающего воздуха до 6—8 г (3—4 л) CO2 и одновременно выделять в окружающий воздух столько же по объёму O2. Высокая эффективность обеспечивается его структурой (толщина листовой пластинки чаще равна 0,1—0,25 мм.) и высокими показателями отношения поверхности (S) к объёму (V) (S/V = 100—200 см2/см3) или к сырой и сухой массе (соответственно 100 и 500 см2/г). Высокая способность Л. к газообмену определяется большой пористостью, наличием в эпидермисе устьиц (до 30 тыс. на 1 см2) и пористым строением мезофилла.
Важное условие высокой фотосинтетической активности — быстрый отток образуемых ассимилятов в др. органы, а в связи с этим — степень развития проводящих систем и прежде всего флоэмы проводящих пучков (жилок). В интенсификации транспорта ассимилятов из Л. в др. органы играет роль у некоторых растений анатомическая структура и, в частности, наличие обкладочных клеток проводящих пучков, в которых сосредоточены хлоропласты и запасаются большие количества крахмала. Многие растения, Л. которых обладают такой структурой (кукуруза, сахарный тростник, сорго и др.), характеризуются особым типом фотосинтеза и его интенсивностью, а также высокой продуктивностью. Непременное условие для фотосинтеза — наличие в хлоропластах ассимилирующих клеток хлорофилла и каротиноидов, содержание которых варьирует у разных растений. При содержании хлорофилла в расчёте на 1 дм2 листовой пластинки 2—4 мг и более количество хлорофилла уже не лимитирует фотосинтез. Л. растений поглощают при этом практически всю энергию входящего в них света с длиной волны от 360 до 720 нм и некоторое количество энергии инфракрасных лучей, пропуская только около 5% энергии (12—15% обычно отражается). При этом в солнечный полдень 1 м2 пластинки Л. может поглотить до 1,5 Мдж (360 ккал) энергии в час. Но в лучшем случае только 5—10% этой энергии превращается в энергию химических связей, остальная превращается в тепло и, нагревая Л., частично уходит в окружающую среду в результате теплоотдачи. При этом не происходит перегрева Л., т.к. около 247 кдж (59 ккал) затрачивается на испарение каждых 100 г воды, которую Л. испаряет в большом кол-ве (в данном примере 1 м2 листовой поверхности, образуя в час при наивысшей освещённости около 4—6 г ассимилятов, может испарять до 500—700 г воды). Однако часто, например в зонах недостаточного водоснабжения, у растений формируются Л., строение которых обеспечивает лучшее снабжение водой, а иногда даже большее её испарение, что увеличивает так называемую сосущую силу. При временных нарушениях водного баланса в Л. активизируются регуляторные системы — движения устьиц, сокращения водоотдачи клетками и т.д., что одновременно ведёт к ослаблению интенсивности газообмена, а следовательно, и фотосинтеза. Сохранению водно-теплового баланса на оптимальном уровне способствуют также восковой налёт на Л., опушённость эпидермиса, сокращение числа устьиц, утолщение Л. в результате сильного развития хорошо оводнённой ткани, превращение Л. в колючки, чешуи и т.п. (функцию фотосинтеза выполняют при этом содержащие хлорофилл стеблевые органы). Редуцированы Л. и у многих растений-паразитов, которые питаются органическими веществами не за счёт фотосинтеза, а добывая их через присоски у растений-хозяев.
Однако не отдельный Л. и не одиночное растение, а их совокупность в естественных или культурных фитоценозах как целостных фотосинтезирующих системах обеспечивает наилучшее использование Л. в процессе фотосинтеза потоков приходящей на поверхность Земли энергии солнечного света, CO2 из воздуха и факторов почвенного плодородия, а следовательно, наивысшую продуктивность. При благоприятных условиях фитоценозы большинства растений практически могут поглощать всю энергию входящего в них света и давать максимальную продукцию, образуя оптимальную площадь Л. — 4—5 м2 на 1 м2 занимаемой ими площади земли. Т. о., получение посевов и насаждений с оптимальной площадью Л. и динамикой её формирования — важнейшая задача повышения продуктивности растений. Эта задача решается по-разному, но на основе общих закономерностей фотосинтетической деятельности Л. растения в фитоценозах. Т. о., через фотосинтез, осуществляемый преимущественно в Л., реализуется космическая функция зелёных растений, которые поглощают солнечную энергию, используя её на биосинтез органического вещества. Через процессы фотосинтеза и дыхания Л. поддерживают баланс O2 и CO2 в воздухе, через транспирацию Л. участвуют в круговороте воды на Земле.
Использование листьев в хозяйственной деятельности человека разнообразно. Л. многих растений употребляют в пищу (салат, щавель, укроп, капуста, лук, петрушка и т.д.), из Л. белены, сенны, наперстянки и многих др. получают лекарственные вещества. Л. др. растений используют на корм с.-х.(сельскохозяйственный) животным в виде сена, сенажа, силоса и свежей травы, Л. шелковицы — на корм для тутового шелкопряда. Из Л. герани, мяты и др. растений получают эфирные масла, используемые в парфюмерной и кондитерской промышленности. Из Л. некоторых растений добывают дубильные вещества. Л. чайного куста и табака служат сырьём в чайной и табачной промышленности. В тропиках Л. многих растений используют для получения грубого волокна, а Л. пальм — как кровельный материал.
Лит.: Тимирязев К. А., Жизнь растения (лекция 5), Избр. соч.(сочинение), т. 1, М., 1957; Серебряков И. Г., Морфология вегетативных органов высших растений, М., 1952; фотосинтезирующие системы высокой продуктивности, Сб., М., 1966; Ничипорович А. А., фотосинтез и теория получения высоких урожаев, М., 1956; Федоров А. А., Кирпичников М. Э., Артюшенко З. Т., Атлас по описательной морфологии высших растений. Лист, М. — Л., 1956; Тахтаджян А. Л., Основы эволюционной морфологии покрытосеменных, М. — Л., 1964; Ботаника, 7 изд., т. 1, М., 1966; Эсау К., Анатомия растений, М., 1969; фотосинтез и использование солнечной энергии, Сб., Л., 1971.
М. Э. Кирпичников, А. А. Ничипорович, А. А. Федоров.