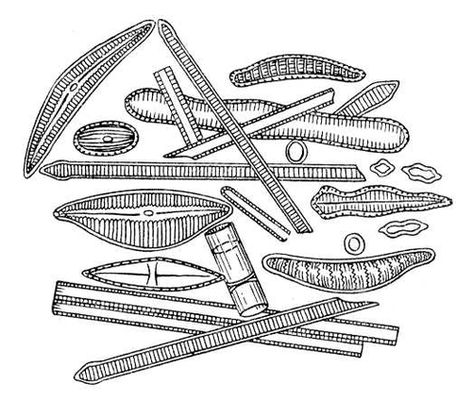
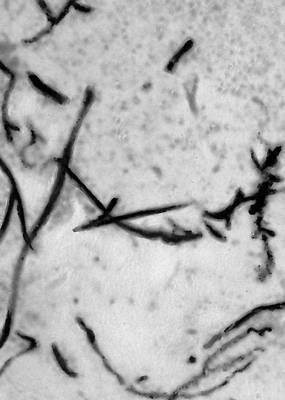
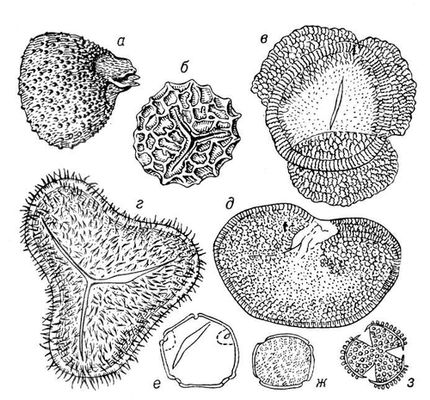
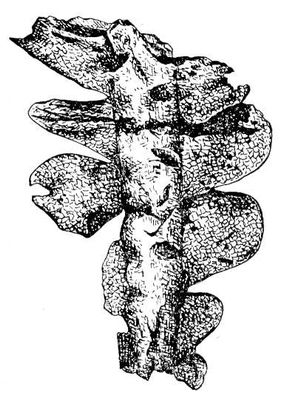
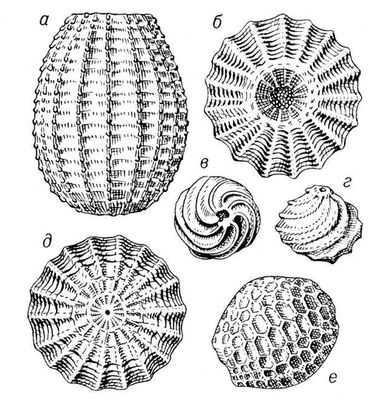
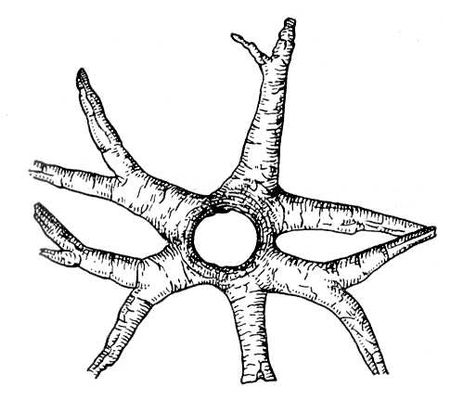
Ископаемые растения, растения прошлых геологических периодов, остатки которых сохранились в отложениях земной коры. Изучение И. р. — предмет палеоботаники. В целом виде сохраняются низшие растения (водоросли и бактерии, рис. 1а—в, 2,3), из высших — редко мхи (рис. 4) и некоторые мелкие формы, от крупных высших растений — только их разрозненные части: стебли, кора, листья, цветки, плоды, семена, споры и пыльцевые зёрна (рис. 5а—д, 6а—б, 7—9, 10а—в). Эта фрагментарность остатков И. р. сильно затрудняет их изучение. Обычно И. р. встречаются в отложениях болот, озёр, рек, а также в известковистых и кремнистых отложениях источников, реже в янтаре и лавах, а также в прибрежно-морских отложениях наряду с ископаемыми животными. Чаще всего в ископаемом состоянии встречаются остатки древесных растений, росших недалеко от водоёмов, а также водные растения и травы, обитавшие в водоёмах и по их берегам. От растений, произраставших вдали от водоёмов, а также растений высокогорных поясов и травянистых равнин в ископаемом состоянии сохранились в основном пыльца и обломки древесины. Таким образом, скопления остатков И. р. могут дать лишь частичное представление о растительном покрове прошлого, преимущественно о древесных формах и о растениях, обитавших вблизи водоёмов.
Степень сохранности остатков зависит и от геологического возраста включающих И. р. пород. Некоторые части И. р. антропогена — древесина, плоды, листья — имеют почти все свойства только что отмерших растений: режутся ножом, в известной мере сохраняют первоначальный цвет и т. п. (см. Ископаемые остатки организмов). Объекты, сохранившие анатомическое строение, исследуют на тонких шлифах или срезах при помощи микроскопа, также изучают и кутикулу, на которой отпечатались клетки эпидермиса, мелкие одноклеточные или колониальные водоросли (диатомовые, синезелёные, жгутиковые), споры и пыльцу растений. Для исследования более крупных остатков пользуются обычно лупой или бинокулярным микроскопом. Основное внимание при изучении листьев обращают на детали их строения — жилкование, форму листа, распределение устьиц, форму клеток эпидермиса листа и т. д.
К И. р. применяют те же методы классификации, что и к современным растениям. Однако многие И. р. принадлежат к ныне вымершим классам и семействам растений. Разрозненность вегетативных частей и органов размножения И. р., изменения, происшедшие с ними, их фрагментарное состояние нередко делают невозможным отнесение тех или иных находок к определённым систематическим группам растений и заставляют временно применять к ним искусственную классификацию, объединяя их по внешнему сходству. При этом различные по своему происхождению растения могут быть отнесены к одному «роду», обычно называемом формальным, и наоборот, части одного и того же растения могут быть отнесены к различным «родам». Например, у лепидодендронов различные формы сохранения их стволов выделяются как «роды» Knorria, Bergeria, Aspidiaria. Своеобразные корнеподобные образования многих лепидофитов (Sigillaria, Bothrodendron и др.) объединяются в «род» Stigmaria; слепки внутренней полости кордаитов называются Artisia. Искусственно классифицируют споры и пыльцу из палеозойских и в значительной мере из мезозойских отложений. К растениям палеогена и особенно неогена, а также антропогена искусственная классификация в СССР почти не применяется.
Изучение И. р. установило присутствие бактерий и одноклеточных водорослей в отложениях архея и протерозоя, т. е. более 3 млрд. лет назад. Наземные растения появились, возможно, в кембрии — ордовике и, несомненно, в силуре. Для начала девонского периода установлено преобладание простейших высших споровых — псилофитов. Почти одновременно возникли такие группы высших споровых растений, как плауновидные, затем прапапоротники, а в конце девона членистостебельные и праголосеменные (археоптерис) и, наконец, птеридоспермы. Последние наряду с папоротниками, плауновидными и членистостебельными приобрели широкое распространение в карбоне, где многие из них были представлены древовидными формами. Хвойные, гинкговые, цикадовые и беннеттитовые появились в пермском или в конце каменноугольного периода, но достигли максимального развития в мезозое. Остатки покрытосеменных растений известны уже из нижнемеловых отложений, однако господствующее положение эти группы растений заняли в позднемеловую эпоху.
Большинство высших споровых и голосеменных растений палеозоя и частично мезозоя принадлежит к вымершим классам, порядкам и семействам растений. Из современного семейства папоротников раньше других появились Marattiaceae, Osmundaceae, Schizaeaceae, затем Cyatheaceae, Dipteridaceae, тогда как Polypodiaceae — гораздо позже. Некоторые роды растений, существовавшие в мезозое, живут до сих пор, например: хвощ Equisetum, папоротники Osmunda, Gleichenia, голосеменное растение гинкго. В начале третичного периода сформировались почти все ныне живущие роды растений. Современные виды появились только в неогене; в плиоцене их число уже значительно. Во 2-й половине третичного и в четвертичном периоде происходили преимущественно образование современных видов, вымирание древних, некогда широко распространённых растений, сохранившихся теперь как реликты (Taxodium, Metasequoia, Sequoia, Ginkgo, Liriodendron, Cercidiphyllum), и особенно географическое перераспределение растительного покрова, принявшего современный вид по окончании ледниковой эпохи.
Эволюция низших споровых растений происходила более замедленными темпами; почти все типы водорослей уже существовали в древнейшие периоды жизни Земли. Только диатомовые и кокколитовые одноклеточные водоросли появились, видимо, в юре. Из девона уже известны разнообразные харовые водоросли (Trochiliscus, Sycidium), в мезозое произошло бурное развитие сифонниковых водорослей, панцири которых образуют мощные скопления в триасовых отложениях Тироля и Ломбардии, а в кайнозое — багряных, или красных, водорослей, участвующих вместе с кораллами в построении рифов. В докембрии были широко распространены рифообразующие колониальные синезелёные водоросли, постройки которых называются строматолитами, а также мельчайшие одноклеточные формы.
И. р. образуют скопления полезных ископаемых, используемых в промышленности (залежи торфа, бурых и каменных углей, горючих сланцев); отложения панцирей различных диатомовых водорослей образуют залежи диатомитов. Изучение И. р. даёт представление о составе и распределении растительного покрова Земли в прошлом и намечает этапы эволюции растений. Состав и распределение флоры в каждой геологической системе и её более дробных подразделениях позволяют (наряду с данными об ископаемых животных и даже при их отсутствии) определять возраст геологических отложений, что имеет особое значение для континентальных образований.
Лит.: Криштофович А. Н., Палеоботаника, 4 изд., Л., 1957; Основы палеонтологии. Водоросли, мохообразные, псилофитовые, плауновидные, членистостебельные, папоротники, М., 1963; Основы палеонтологии. Голосеменные и покрытосеменные, М., 1963; Палеопалинология, под ред. И. М. Покровской, т. 1—3, Л., 1966; Палеозойские и мезозойские флоры Евразии и фитогеография этого времени, М., 1970; Съюорд А. Ч., Века и растения. Обзор растительности прошлых геологических периодов, пер.(перевод) [с англ.(английский)], Л.—М., 1936; Seward А. С., Fossil plants, v. 1—4, Camb., 1898—1919; Gothan W., Weyland Н., Lehrbuch der Paläobotanik, 2 Aufl., B., 1964.