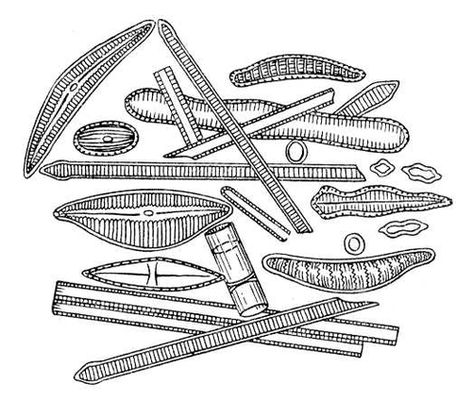
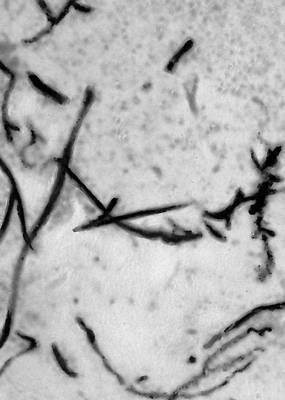
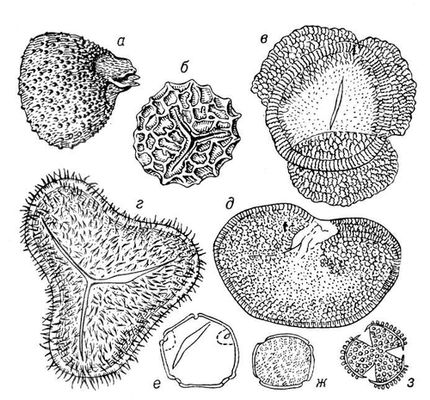
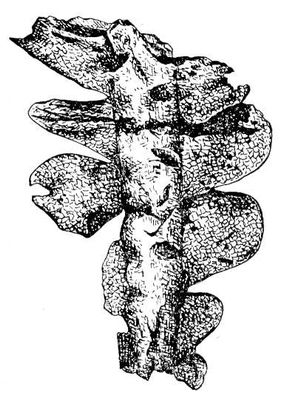
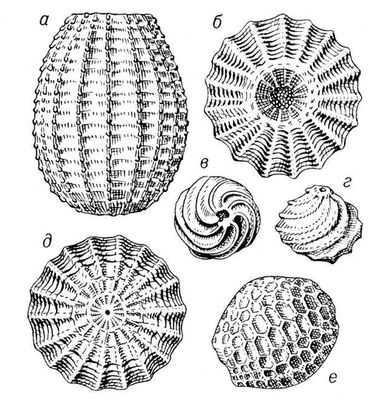
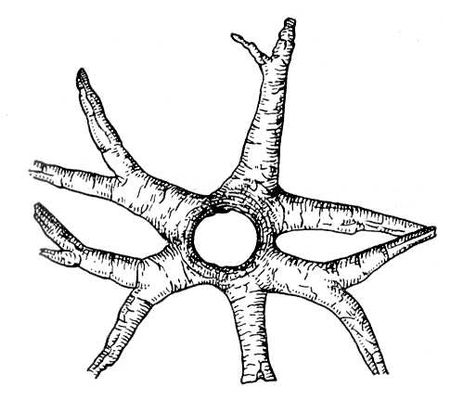
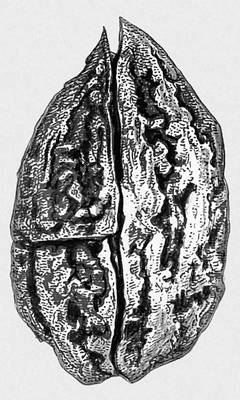
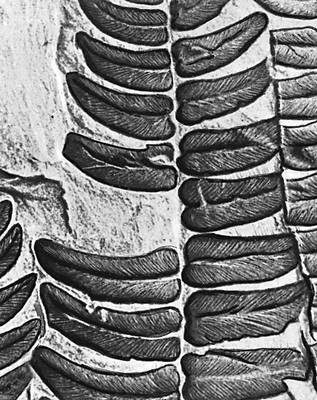
Викопні рослини, рослини минулих геологічних періодів, залишки яких збереглися у відкладеннях земної кори. Вивчення І. р. — предмет палеоботаніки . У цілому вигляді зберігаються нижчі рослини (водорості і бактерії, мал. 1а—в , 2 , 3 ), з вищих — рідко мохи ( мал. 4 ) і деякі дрібні форми, від крупних вищих рослин — лише їх розрізнені частини: стебла, кора, листя, квітки, плоди, насіння, спори і пилкові зерна ( мал. 5а—д, 6а—б, 7—9, 10а—в ). Ця фрагментарність залишків І. р. сильно утрудняє їх вивчення. Зазвичай І. р. зустрічаються у відкладеннях боліт, озер, річок, а також в ізвестковістих і крем'янистих відкладеннях джерел, рідше в янтарі і лавах, а також в прибережно-морських відкладеннях поряд з копалинами тваринами . Найчастіше у викопному стані зустрічаються залишки деревних рослин, що зростали недалеко від водоймищ, а також водні рослини і трави, що мешкали у водоймищах і по їх берегах. Від рослин, що виростали далеко від водоймищ, а також рослин високогірних поясів і трав'янистих рівнин у викопному стані збереглися в основному пилок і уламки деревини. Таким чином, скупчення залишків І. р. можуть дати лише часткове уявлення про рослинний покрив минулого, переважно про деревні форми і про рослини, водоймищ, що мешкали поблизу.
Міра збереження залишків залежить і від геологічного віку тих, що включають І. р. порід. Деякі частини І. р. антропогена — деревина, плоди, листя — мають майже всі властивості тільки що відмерлих рослин: ріжуться ножем, певною мірою зберігають первинний колір і тому подібне (див. Викопні залишки організмів ). Об'єкти, що зберегли анатомічну будову, досліджують на тонких шліфах або зрізах за допомогою мікроскопа, також вивчають і кутикулу, на якій віддрукувалися клітки епідермісу, дрібні одноклітинні або колоніальні водорості (діатомові, синезелениє, жгутикові), спори і пилок рослин. Для дослідження крупніших залишків користуються зазвичай лупою або бінокулярним мікроскопом. Основна увага при вивченні листя обертають на деталі їх будови — жилкування, форму аркуша, розподіл устьіц, форму кліток епідермісу аркуша і так далі
До І. р. застосовують ті ж методи класифікації, що і до сучасних рослин. Проте багато І. р. належать до нині вимерлих класів і сімейств рослин. Розрізненість вегетативних частин і органів розмноження І. р., зміни, що сталися з ними, їх фрагментарний стан незрідка роблять неможливим віднесення тих або інших знахідок до певних систематичних груп рослин і заставляють тимчасово застосовувати до них штучну класифікацію, об'єднуючи їх по зовнішній схожості. При цьому різні по своєму походженню рослини можуть бути віднесені до одного «роду», зазвичай званому формальним, і навпаки, частини однієї і тієї ж рослини можуть бути віднесені до різних «пологів». Наприклад, в лепідодендронов різні форми збереження їх стволів виділяються як «пологи» Knorria Bergeria, Aspidiaria. Своєрідні коренеподібні утворення багатьох лепідофітов (Sigillaria, Bothrodendron і ін.) об'єднуються в «рід» Stigmaria; зліпки внутрішньої порожнини кордаїтов називаються Artisia. Штучно класифікують спори і пилок з палеозойських і значною мірою з мезозойських відкладень. До рослин палеогену і особливо неогена, а також антропогена штучна класифікація в СРСР майже не застосовується.
Вивчення І. р. встановило присутність бактерій і одноклітинних водоростей у відкладеннях архея і протерозоя, тобто більше 3 млрд. років назад. Наземні рослини з'явилися, можливо, в кембрії — ордовіке і, поза сумнівом, в силурі. Для початку девонського періоду встановлено переважання простих вищих спорових — псилофітов . Майже одночасно виникли такі групи вищих спорових рослин, як плауновідниє, потім прапапоротникі, а в кінці девона членістостебельниє і праголосеменниє (археоптеріс) і, нарешті, птерідосперми. Останні поряд з папоротями, плауновіднимі і членістостебельнимі придбали широке поширення в карбоні, де багато хто з них був представлений деревовидними формами. Хвойні, гинкговиє, цикадовиє і беннеттітовиє з'явилися в пермському або в кінці кам'яновугільного періоду, але досягли максимального розвитку в мезозої. Залишки покритосеменних рослин відомі вже з ніжнемелових відкладень, проте пануюче положення ці групи рослин зайняли в позднемеловую епоху.
Більшість вищих спорових і голосеменних рослин палеозою і частково мезозою належить до вимерлих класів, порядків і сімейств рослин. З сучасного сімейства папоротей раніше інших з'явилися Marattiaceae, Osmundaceae, Schizaeaceae, потім Cyatheaceae, Dipteridaceae, тоді як Polypodiaceae — набагато пізніше. Деякі пологи рослин що існували в мезозої, живуть до цих пір, наприклад: хвощ Equisetum, папороті Osmunda, Gleichenia, голосеменноє рослина гинкго. На початку третинного періоду сформувалися майже всі пологи рослин, що нині живуть. Сучасні види з'явилися лише в неогене; у пліоцені їх число вже значно. У 2-ій половині третинного і в четвертинному періоді відбувалися переважно утворення сучасних видів, вимирання древніх, колись широко поширених рослин що збереглися тепер як релікти (Taxodium, Metasequoia, Sequoia, Ginkgo, Liriodendron, Cercidiphyllum), і особливо географічний перерозподіл рослинного покриву, що набрав сучасного вигляду після закінчення льодовикової епохи.
Еволюція нижчих спорових рослин відбувалася більш сповільненими темпами; майже всі типи водоростей вже існували в прадавні періоди життя Землі. Лише діатомові і кокколітовиє одноклітинні водорості з'явилися мабуть, в юрі. З девона вже відомі всілякі харовиє водорості (Trochiliscus, Sycidium), в мезозої стався бурхливий розвиток сифонникових водоростей, панцирі яких утворюють потужні скупчення у відкладеннях тріасів Тироля і Ломбардії, а в кайнозої — багряних, або червоних, водоростей, що беруть участь разом з коралами в побудові рифів. У докембрії були широко поширені ріфообразующие колоніальні синезелениє водорості, споруди яких називаються строматолітамі а також найдрібніші одноклітинні форми.
І. р. утворюють скупчення корисних копалини, використовуваних в промисловості (поклади торфу, бурого і кам'яного вугілля, горючих сланців); відкладення панцирів різних діатомових водоростей утворюють поклади діатомітов. Вивчення І. р. дає уявлення про склад і розподіл рослинного покриву Землі у минулому і намічає етапи еволюції рослин. Склад і розподіл флори в кожній геологічній системі і її більш дробових підрозділах дозволяють (поряд з даними про копалини тварин і навіть при їх відсутності) визначати вік геологічних відкладень, що має особливе значення для континентальних утворень.
Літ.: Кріштофовіч А. Н., Палеоботаніка, 4 видавництва, Л., 1957; Основи палеонтології. Водорості, мохоподібні, псилофітовиє, плауновідниє, членістостебельниє, папороті, М., 1963; Основи палеонтології. Голосеменниє і покритосеменниє М., 1963; Палеопалінология, під ред. І. М. Покровськой, т. 1—3, Л., 1966; Палеозойські і мезозойські флори Євразії і фітогеографія цього часу, М., 1970; С'юорд А. Ч., Століття і рослини. Огляд рослинності минулих геологічних періодів, пер.(переведення) [з англ.(англійський)], Л.—М., 1936; Seward А. С., Fossil plants, v. 1—4, Camb., 1898—1919; Gothan W., Weyland Н., Lehrbuch der Paläobotanik, 2 Aufl., B., 1964.