Імуногенетика, комплексна наукова дисципліна, що поєднує методи імунології, молекулярної біології і генетики для вивчення спадкових чинників імунітету, внутрівидової різноманітності і спадкоємства тканинних антигенів, генетичних і популяцій аспектів взаємин макро- і мікроорганізму і тканинній несумісності . Почало І. поклали роботи німецьких учених П. Ерліха і Ю. Моргенрота, що виявили на початку 20 ст групи крові у кіз, і відкриття К. Ландштейнером груп крові у людини. Термін «І.» запропонований американським ученим М. Ірвіном в 1930.
Індивідуальна і видова стійкість рослин і тварин до бактерійним і вірусним інфекціям забезпечується складною багатоступінчастою системою захисних сил організму. У боротьбі між захисними силами і інфекційними агентами «перевага» часто залишається на стороні останніх, оскільки мікроорганізми швидко розмножуються, утворюючи багатомільйонні популяції, в яких рано чи пізно виникають форми мутантів з агресивнішими властивостями, ніж у вихідного штаму. Ймовірно, як у відповідь захисний засіб на певному етапі еволюції хребетних тварин виникла система адаптивного імунітету (антітелообразованіє) — найбільш потужна лінія оборони організму, особливо при повторних контактах з інфекційними агентами. Здатність (або нездатність) виробляти антитіла — спадкову ознаку. Генетична регуляція біосинтезу антитіл має характерні особливості. Так, утворення одного поліпептидного ланцюга молекули антитіла контролюється двома різними генами . Один з них контролює утворення частини ланцюга, що бере участь в побудові активного центру ; будова цієї частини різна в антитіл різної специфічності. Інший ген контролює утворення частини ланцюга, будова якого однаково в антитіл, що відносяться до даного класу імуноглобулінів .
Окрім групових антигенів, існують успадковані їх варіанти, специфічні для окремих типів кліток, наприклад для лейкоцитів. Відмінності в будові лейкоцитарних антигенів у донора і реципієнта — одна з причин несумісності при пересадці органів і тканин. Спадкові внутрівидові відмінності в будові багатьох білків сироватки крові (альбумін, трансферріни і ін.) контролюються, як правило, алельними генами, причому частота кожній аллелі в популяції висока (20% і вище), що вказує на «тиск» природного відбору. Одне з найважливіших завдань І. — встановлення чинників, що обумовлюють поширення в популяціях нових аллелей. Таким чинником може служити схожість в будові антигенів у хвороботворних мікроорганізмів і макроорганізму. Тварини в нормі не виробляють антитіл до власних антигенів, тому схожість в антигенній будові між яким-небудь компонентом мікробної клітки і тією або іншою молекулою макроорганізму приведе до того, що останній не зможе синтезувати антитіла, що знешкоджують даний вигляд мікроба. У зв'язку з цим знижуються захисні сили макроорганізму. Тому відбір підхоплюватиме появу видозмінених молекул білків (або полісахаридів), підвищуючи тим самим імунну стійкість організму. Поширення в популяції нових аллелей може відбуватися також і в тих випадках, коли в результаті мутації відповідного гена молекула макроорганізму змінюється так, що ферментативні системи мікроба вже не можуть її використовувати як субстрат. Інколи для цього досить заміни однієї амінокислоти в поліпептидному ланцюзі, як це має місце в деяких форм мутантів гемоглобіну . Такі форми поширилися в районах земної кулі, де висока захворюваність малярією: носії гемоглобіну мутанта не хворіють на малярію, оскільки малярійний плазмодій нездібний використовувати його як субстрат. У ряді випадків поширюються мутації, які змінюють біохімію клітки або органу в цілому і тим самим порушують пристосованість паразита. Мабуть, існують та інші механізми спадкового імунітету, завдяки яким досягається спадкова гетерогенність вигляду-господаря, що перешкоджає поширенню паразитичного штаму мікроорганізму.
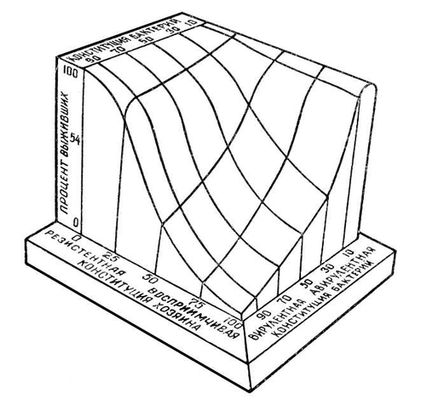
Таким чином, міра природної стійкості до захворювання тварин даного вигляду визначається багатьма чинниками, сумарно відображаючи особливості конституції і тварини, і збудника захворювання. Тривимірна модель цих взаємин представлена на мал. , де показано, що відсоток особин, що вижили після інфекції, залежить як від спадкової стійкості організму до збудника захворювання, так і від вірулентності останнього.
Спадкова стійкість до захворювань, як правило, специфічна, оскільки фізіологічні основи стійкості до різних захворювань зазвичай неоднакові. Так, африканська худоба зебу, що прекрасно переносить жару і стійкий до туберкульозу дуже чутливий до тріпаносомозу; лінія білих леггорнів, стійка до моноцитозу курнув, чутлива до курячого лейкозу; лінії мишей, стійкі до мишачого тифу, надзвичайно сприйнятливі до вірусу помилкового сказу. З прадавніх часів генетична стійкість окремих особин, порід, рас і так далі до захворювань служила передумовою для селекції . Так були виведені вівці породи ромні-марш, стійкі до тріхостронгилідам, раса кроликів стійка до міксоматозу, і медоносні бджоли, стійкі до американського гнильця. Природний відбір на стійкість існував і серед людей. Так, після відкриття Нового Світла виявилось, що індійці Північної Америки чутливіші до кору і вітряної віспи, чим європейці, для яких ці захворювання були звичні і легко переносими.
В основі генетичної стійкості до захворювань лежать всілякі механізми, у тому числі і неімунологічні. Білі леггорни, наприклад, стійкі до білого проносу тому, що мають досконалішу терморегуляцію; стійкість худоби зебу до кліщових захворювань обумовлена товщою шкірою і особливостями шкірних виділень, які відлякують кліщів. Чутливість до віспи в осіб з групами крові А і AB пов'язана із спільністю антигена А людини і антигенів вірусу віспи. Тому особи з групами крові В і Про(Н) легше переносять віспу.
Перенесення генетичних вистав в область імунології дозволило радянському ученому В. П. Ефроїмсону сформулювати еволюційно-генетичну концепцію імуногенезу, що пояснює внутрівидову антигенну різноманітність і гетерогенність антитіл по специфічності. Кожна здорова зріла в імунологічному відношенні особина здібна до імунної відповіді на тканинні антигени особини з іншим генотипом. Таким чином, тканинна несумісність — універсальна біологічна закономірність. Лише однояйцеві близнята і тварини одній чистій лінії не розділені бар'єром тканинної несумісності, вираженість якої залежить від міри відмінності генотипів донора і реципієнта. Для успішних пересадок органів і тканин, переливань крові і клітин кісткового мозку дуже поважно понизити до мінімуму величину цієї відмінності шляхом підбору сумісного донора. Вивчення клітинних антигенів, їх спадкоємства і різноманітності, їх виявлення (тіпірованіє) — це ті розділи І., які особливо важливі для трансплантології, трансфузіологиі, імуногематології і клінічній імунології. Див. також Імунологія .
Літ.: Медведев Н. Н., Лінійні миші, Л., 1964; Хатт Ф., Генетика тварин. пер.(переведення) з англ.(англійський), М., 1969; Ефроїмсон Ст П., Імуногенетика, М., 1971; Hildemann W. Н., Immunogenetics, San Francisco, 1970.