Иммуногенетика, комплексная научная дисциплина, сочетающая методы иммунологии, молекулярной биологии и генетики для изучения наследственных факторов иммунитета, внутривидового разнообразия и наследования тканевых антигенов, генетических и популяционных аспектов взаимоотношений макро- и микроорганизма и тканевой несовместимости. Начало И. положили работы немецких учёных П. Эрлиха и Ю. Моргенрота, обнаруживших в начале 20 в. группы крови у коз, и открытие К. Ландштейнером групп крови у человека. Термин «И.» предложен американским учёным М. Ирвином в 1930.
Индивидуальная и видовая устойчивость растений и животных к бактериальным и вирусным инфекциям обеспечивается сложной многоступенчатой системой защитных сил организма. В борьбе между защитными силами и инфекционными агентами «преимущество» часто остаётся на стороне последних, так как микроорганизмы быстро размножаются, образуя многомиллионные популяции, в которых рано или поздно возникают мутантные формы с более агрессивными свойствами, чем у исходного штамма. Вероятно, как ответное защитное средство на определённом этапе эволюции позвоночных животных возникла система адаптивного иммунитета (антителообразование) — наиболее мощная линия обороны организма, особенно при повторных контактах с инфекционными агентами. Способность (или неспособность) вырабатывать антитела — наследственный признак. Генетическая регуляция биосинтеза антител имеет характерные особенности. Так, образование одной полипептидной цепи молекулы антитела контролируется двумя разными генами. Один из них контролирует образование части цепи, участвующей в построении активного центра; строение этой части различно у антител разной специфичности. Другой ген контролирует образование части цепи, строение которой одинаково у антител, относящихся к данному классу иммуноглобулинов.
Помимо групповых антигенов, существуют наследуемые их варианты, специфичные для отдельных типов клеток, например для лейкоцитов. Различия в строении лейкоцитарных антигенов у донора и реципиента — одна из причин несовместимости при пересадке органов и тканей. Наследственные внутривидовые различия в строении многих белков сыворотки крови (альбумины, трансферрины и др.) контролируются, как правило, аллельными генами, причём частота каждой аллели в популяции высока (20% и выше), что указывает на «давление» естественного отбора. Одна из важнейших задач И. — установление факторов, обусловливающих распространение в популяциях новых аллелей. Таким фактором может служить сходство в строении антигенов у болезнетворных микроорганизмов и макроорганизма. Животные в норме не вырабатывают антител к собственным антигенам, поэтому сходство в антигенном строении между каким-либо компонентом микробной клетки и той или иной молекулой макроорганизма приведёт к тому, что последний не сможет синтезировать антитела, обезвреживающие данный вид микроба. В связи с этим снижаются защитные силы макроорганизма. Поэтому отбор будет подхватывать появление видоизменённых молекул белков (или полисахаридов), повышая тем самым иммунную устойчивость организма. Распространение в популяции новых аллелей может происходить также и в тех случаях, когда в результате мутации соответствующего гена молекула макроорганизма изменяется так, что ферментативные системы микроба уже не могут её использовать в качестве субстрата. Иногда для этого достаточно замены одной аминокислоты в полипептидной цепи, как это имеет место у некоторых мутантных форм гемоглобина. Такие формы распространились в районах земного шара, где высока заболеваемость малярией: носители мутантного гемоглобина не болеют малярией, так как малярийный плазмодий неспособен использовать его в качестве субстрата. В ряде случаев распространяются мутации, которые изменяют биохимию клетки или органа в целом и тем самым нарушают приспособленность паразита. По-видимому, существуют и другие механизмы наследственного иммунитета, благодаря которым достигается наследственная гетерогенность вида-хозяина, препятствующая распространению паразитического штамма микроорганизма.
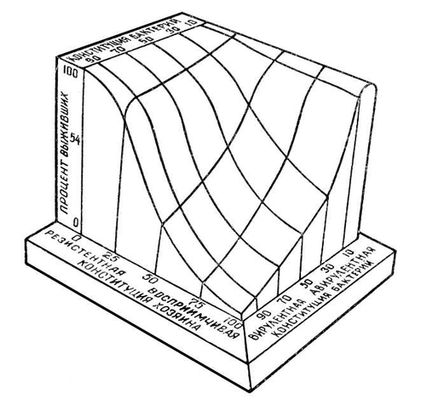
Таким образом, степень естественной устойчивости к заболеванию животных данного вида определяется многими факторами, суммарно отражая особенности конституции и животного, и возбудителя заболевания. Трёхмерная модель этих взаимоотношений представлена на рис., где показано, что процент особей, выживших после инфекции, зависит как от наследственной устойчивости организма к возбудителю заболевания, так и от вирулентности последнего.
Наследственная устойчивость к заболеваниям, как правило, специфична, так как физиологические основы устойчивости к разным заболеваниям обычно неодинаковы. Так, африканский скот зебу, прекрасно переносящий жару и устойчивый к туберкулёзу очень чувствителен к трипаносомозу; линия белых леггорнов, устойчивая к моноцитозу кур, чувствительна к куриному лейкозу; линии мышей, устойчивые к мышиному тифу, чрезвычайно восприимчивы к вирусу ложного бешенства. С древнейших времён генетическая устойчивость отдельных особей, пород, рас и т. д. к заболеваниям служила предпосылкой для селекции. Так были выведены овцы породы ромни-марш, устойчивые к трихостронгилидам, раса кроликов, устойчивая к миксоматозу, и медоносные пчёлы, устойчивые к американскому гнильцу. Естественный отбор на устойчивость существовал и среди людей. Так, после открытия Нового Света оказалось, что индейцы Северной Америки более чувствительны к кори и ветряной оспе, чем европейцы, для которых эти заболевания были привычны и легко переносимы.
В основе генетической устойчивости к заболеваниям лежат разнообразные механизмы, в том числе и неиммунологические. Белые леггорны, например, устойчивы к белому поносу потому, что имеют более совершенную терморегуляцию; устойчивость скота зебу к клещевым заболеваниям обусловлена более толстой кожей и особенностями кожных выделений, которые отпугивают клещей. Чувствительность к оспе у лиц с группами крови А и AB связана с общностью антигена А человека и антигенов вируса оспы. Поэтому лица с группами крови В и О(Н) легче переносят оспу.
Перенесение генетических представлений в область иммунологии позволило советскому учёному В. П. Эфроимсону сформулировать эволюционно-генетическую концепцию иммуногенеза, объясняющую внутривидовое антигенное разнообразие и гетерогенность антител по специфичности. Каждая здоровая зрелая в иммунологическом отношении особь способна к иммунному ответу на тканевые антигены особи с другим генотипом. Таким образом, тканевая несовместимость — универсальная биологическая закономерность. Лишь однояйцевые близнецы и животные одной чистой линии не разделены барьером тканевой несовместимости, выраженность которой зависит от степени несходства генотипов донора и реципиента. Для успешных пересадок органов и тканей, переливаний крови и клеток костного мозга очень важно снизить до минимума величину этого несходства путём подбора совместимого донора. Изучение клеточных антигенов, их наследования и разнообразия, их обнаружение (типирование) — это те разделы И. , которые особенно важны для трансплантологии, трансфузиологии, иммуногематологии и клинической иммунологии. См. также Иммунология.
Лит.: Медведев Н. Н., Линейные мыши, Л., 1964; Хатт Ф., Генетика животных. пер.(перевод) с англ.(английский), М., 1969; Эфроимсон В. П., Иммуногенетика, М., 1971; Hildemann W. Н., Immunogenetics, San Francisco, 1970.