М'язи, мускулатура скелетна і внутрішніх органів (вісцелярна), забезпечуюча у тварин і людини виконання ряду найважливіших фізіологічних функцій: переміщення тіла або окремих його частин в просторі, кровообіг, дихання, пересування харчової кашки в травних органах, підтримка тонусу судин, виділення екськрементов і т. д. Скоротлива функція всіх типів М. обумовлена перетворенням в м'язових волокнах хімічної енергії певних біохімічних процесів в механічну роботу. Проте скорочення скелетних М. і мускулатури внутрішніх органів — лише окремий випадок загальнішої закономірності — механо-хімічній активності живих структур. Мабуть, в основі самих різних проявів цієї активності — скорочення хвоста сперматозоїда, рухи вій інфузорій, розбіжності хромосом під час мітоза, уприскування в бактерію фагової ДНК(дезоксирибонуклеїнова кислота) і т. д. — лежить один і той же молекулярний механізм, пов'язаний з можливістю зміни конформації або взаємного розташування фіблярних структур контрактильних білків.
Типи м'язів . Морфологи розрізняють 2 основних типа М.: поперечнополосатиє м'язи і гладкі м'язи . До перших відноситься вся скелетна мускулатура хребетних тварин і людини, що забезпечує можливість виконання довільних рухів, М. мови, верхній третині стравоходу і деякі ін., М. серця (міокард), що має свої особливості (склад білків, характер скорочення і ін.), а також М. членистоногих і деяких ін. безхребетних. До гладких М. належить велика частина мускулатури безхребетних тваринних і м'язові шари внутрішніх органів і стінок кровоносних судин хребетних тварин і людини, що забезпечують можливість виконання ряду найважливіших фізіологічних функцій. Деякі гістологи, що вивчають М. безхребетних, виділяють і 3-го типа М. — з подвійною косою покресленою (див. М'язова тканина ).
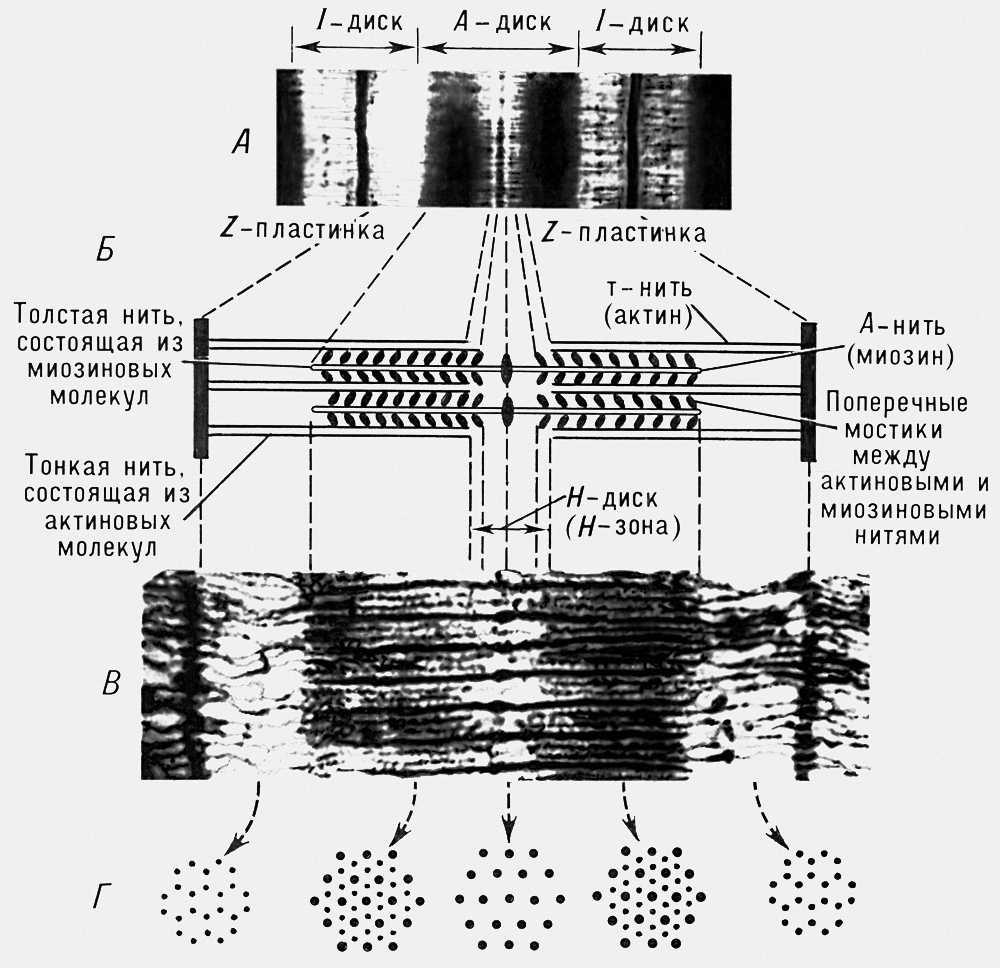
Структурними елементами всіх типів М. є м'язові волокна ( мал. 1 ). Поперечнополосатиє м'язові волокна в скелетних М. утворюють пучки, сполучені один з одним прошарками сполучної тканини. Своїми кінцями м'язові волокна сплітаються з сухожильними волокнами, через посредство яких м'язова тяга передається на кісті скелета. Волокнами поперечнополосатих М. є гігантські багатоядерні клітки, діаметр яких варіює від 10 до 100 мкм , а довжина часто відповідає довжині М., досягаючи, наприклад, в деяких М. людини 12 см . Волокно покрите еластичною оболонкою — сарколемою і складається з саркоплазми, структурними елементами якої є такі органоїди, як мітохондрії, рибосоми, трубочки і бульбашки саркоплазматичної мережі і так звані Т-системі ( мал. 2 ), різні включення і т. д. У саркоплазмі зазвичай у формі пучків розташована безліч ниткоподібних утворень товщиною від 0,5 до декількох мкм — міофібрил, що володіють, як і все волокно в цілому, поперечною покресленою. Кожна міофібрила розділена на декілька сотів ділянок завдовжки 2,5—3 мкм , званих саркомерами. Кожен саркомер, у свою чергу, складається з ділянок, що чергуються, — дисків, що володіють неоднаковою оптичною щільністю і що додають міофібрилам і м'язовому волокну в цілому характерну поперечну покреслену, що чітко виявляється при спостереженні у фазовоконтрастном мікроскопі. Темніші диски володіють здатністю до подвійного променезаломлення і називаються анізотропними, або дисками А . Світліші диски не володіють цією здатністю і називаються ізотропними, або дисками I . Середню частину диска А займає зона слабкішого подвійного променезаломлення — зона Н . Диск I ділиться на 2 рівних частини темною z-пластінкою, що відмежовує один саркомер від іншого. У кожному саркомері є два типи ниток (філаментов), що складаються з м'язових білків: товсті міозіновиє і тонкі — актіновиє ( мал. 3 ). Декілька іншу структуру мають гладкі м'язові волокна. Вони є веретеноподібними одноядерними клітками, позбавленими поперечній покресленій. Довжина їх зазвичай досягає 50—250 мкм (у матці — до 500 мкм ), ширина — 4—8 мкм ; міофіламенти в них зазвичай не об'єднані у відособлені міофібрили, а розташовані по довжині волокна у вигляді безлічі одиночних актінових ниток ( мал. 4 ). Впорядкована система міозінових ниток в гладком'язових клітках відсутня. У гладкій мускулатурі молюсків найбільш важливу роль в здійсненні замикальної функції грають, мабуть, параміозіновиє волокна (тропоміозін А).
Хімічний склад м'язів вагається залежно від вигляду і віку тварини, типа і функціонального стану М. і ряду ін. чинників. Основні речовини, що входять до складу поперечнополосатих М. людини і тварин, і їх вміст (у % до сирої маси) представлені нижчим:
В середньому близько 75% сирої маси М. складає вода. Основна кількість щільних речовин припадає на частку білків. Розрізняють білки міофібрилярні (скоротливі) — міозин, актин і їх комплекс — актоміозін, тропоміозін і ряд так званих мінорних білків (а і b-актініни, тропонін і ін.), і саркоплазматичні — глобуліни X, міогени, дихальні пігменти, зокрема міоглобін нуклеопротєїди і ферменти, що беруть участь в процесах обміну речовин в М. Із ін. з'єднань найважливішими є екстрактні, такі, що беруть участь в обміні речовин і здійсненні скоротливої функції М.: АТФ, фосфокреатин, карнозин, анзерін і др.; фосфоліпіди, що грають важливу роль в утворенні клітинних мікроструктур і в обмінних процесах; безазотисті речовини: глікоген і продукти його розпаду (глюкоза, молочна кислота і ін.), нейтральні жири, холестерин і др.; мінеральні речовини — солі До, Na, Ca, Mg. Гладкі м'язи істотно відрізняються по хімічному складу від поперечнополосатих (нижчий вміст контрактальних білків — актоміозіна, макроергічних з'єднань, діпептідов і ін.).
Функціональні особливості поперечнополосатих м'язів. Поперечнополосатиє М. багато забезпечені різними нервами, за допомогою яких здійснюється регуляція м'язової діяльності з боку нервових центрів. Найважливіші з них: рухові нерви, провідні к М. імпульси, що викликають її збудження і скорочення; чутливі нерви, по яких від М. до нервових центрів поступає інформація про її стан, і, нарешті, адаптаційно-трофічні волокна симпатичної нервової системи, що впливають на обмін речовин і стомлення М., що уповільнюють розвиток (див. Адаптаційно-трофічна функція ).
Кожна гілочка рухового нерва, що іннервує цілу групу м'язових волокон, створюючих так звану моторну одиницю, доходить до окремого м'язового волокна. Всі м'язові волокна, що входять до складу такої одиниці, скорочуються при збудженні практично одночасно. Під впливом нервового імпульсу в закінченнях рухового нерва вивільняється медіатор — ацетілхолін, що взаємодіє з холінорецептором постсинаптичної мембрани (див. Синапси ). В результаті цього відбувається підвищення проникності мембрани для іонів Na і До, що, у свою чергу, обумовлює її деполяризацію (поява постсинаптичного потенціалу). Після цього на сусідніх ділянках мембрани м'язового волокна виникає хвиля збудження (хвиля електронегативності), яка поширюється по скелетному м'язовому волокну зазвичай з швидкістю декілька м-код в 1 сік . В результаті збудження М. змінює свої еластичні властивості. Якщо точки прикріплення М. не фіксовані непорушно, відбувається її укорочення (скорочення). При цьому М. виробляє певну механічну роботу. Якщо точки прикріплення М. нерухомі, в ній розвивається напруга. Між виникненням збудження і появою хвилі скорочення або хвилі напруги протікає деякий час, зване латентним періодом. Скорочення М. супроводиться виділенням тепла, яке продовжується протягом певного часу і після їх розслаблення.
В М. ссавців і людини встановлено існування «повільних» м'язових волокон (до них належать «червоні», такі, що містять дихальний пігмент міоглобін) і «швидких» («білих», таких, що не мають міоглобіну), таких, що розрізняються швидкістю проведення хвилі скорочення і її тривалістю. У «повільних» волокнах ссавцям тривалість хвилі скорочення приблизно в 5 разів більше, а швидкість проведення в 2 рази менша, ніж в «швидких» волокнах. Майже всі скелетні М. відносяться до змішаного типа, тобто містять як «швидкі», так і «повільні» волокна. Залежно від характеру роздратування виникає або одіночноє — фазне — скорочення м'язових волокон, або тривале — тетанічне. Тетанус виникає в разі вступу у М. серії роздратувань з такою частотою, при якій кожне подальше роздратування ще застає М. в стані скорочення, унаслідок чого відбувається підсумовування скоротливих хвиль. Н. Е. Введенський встановив, що збільшення частоти роздратувань викликає зростання тетануса, але лише до відомої межі, званої їм «оптимумом». Подальше почастішання роздратувань зменшує тетанічне скорочення (песимум ). Розвиток тетануса має велике значення при скороченні «повільних» м'язових волокон. У М. з переважанням «швидких» волокон максимальне скорочення — зазвичай результат суммациі скорочень всіх моторних одиниць, в які нервові імпульси поступають, як правило, неодночасно, асихронно.
В поперечнополосатих М. встановлено також існування так званих чисто тонічних волокон, які особливо широко представлені в М. земноводних і плазуючих. Тонічні волокна беруть участь в підтримці мишечного, що не «стомлює», тонусу . Тонічним скороченням називається злите скорочення, що повільно розвивається, здатне тривало підтримуватися без значних енергетичних витрат і що виражається в протидії, що не «стомлює», зовнішнім силам, прагнучим розтягнути м'язовий орган. Тонічні волокна реагують на нервовий імпульс хвилею скорочення лише локально (у місці роздратування). Проте, завдяки великому числу кінцевих рухових бляшок тонічне волокно може збуджуватися і скорочуватися все цілком. Скорочення таких волокон розвивається настільки повільно, що вже при вельми малих частотах роздратування окремі хвилі скорочення накладаються один на одного і зливаються в укорочення, що тривало підтримується. Тривала протидія тонічних волокон, а також повільних фазних волокон розтягуючим зусиллям забезпечується не лише пружною напругою, але і зростанням в'язкості м'язових білків.
Для характеристики скоротливої функції М. користуються поняттям «Абсолютної сили», яка є величиною, пропорційною перетину М., направленою перпендикулярно її волокнам, і виражається в кг/см 2 . Так, наприклад, абсолютна сила двоголовою М. людини рівна 11,4, литковою — 5,9 кг/см 2 .
Систематична посилена робота М. (тренування) збільшує їх масу, силу і працездатність. Проте надмірна робота приводить до розвитку стомлення, тобто до падіння працездатності М. Бездеятельность М. веде до їх атрофії.
Функціональні особливості гладких м'язів. Гладкі М. внутрішніх органів по характеру іннервації, збудження і скорочення істотно відрізняються від скелетних М. Волни збудження і скорочення протікають в гладких М. в дуже сповільненому темпі. Розвиток стану тонусу гладких М., що не «стомлює», зв'язаний, як і в тонічних скелетних волокнах, із сповільненістю скоротливих хвиль, що зливаються один з одним навіть при рідких ритмічних роздратуваннях. Для гладких М. характерна також здатність до автоматизму, тобто до діяльності, не пов'язаної зі вступом в М. нервових імпульсів з центральної нервової системи. Встановлено, що здібністю до ритмічного мимовільного збудження і скорочення володіють не лише нервові клітини, наявні в гладких М., але і самі гладком'язові клітки.
Своєрідність скоротливої функції гладких М. хребетних тварин визначається не лише особливостями їх іннервації і гістологічної будови, але і специфікою їх хімічного складу: нижчим вмістом контрактильних білків (актоміозіна), макроергічних з'єднань, зокрема АТФ, низькою АТФ-АЗНОЙ активністю міозину, наявністю в них водорозчинній модифікації актоміозіна — тоноактоміозіна і т. д.
Істотне значення для організму має здатність гладких м'язів змінювати довжину без підвищення напруги (наповнення порожнистих органів, наприклад сечового міхура, шлунку і ін.).
І. І. Іванов.
Скелетні м'язи людини, різні формою, величині, положенію, складають понад 40% маси його тіла. При скороченні відбувається укорочення М., яке може досягати 60% їх довжин; чим довше за М. (щонайдовша М. тіла кравецька досягає 50 см ), тим більше розмах русі. Скорочення куполоподібної М. (наприклад, діафрагми) обумовлює її сплощення, скорочення кільцеподібних М. (сфінктерів) супроводиться звуженням або закриттям отвору. М. радіального напряму, навпаки, викликають при скороченні розширення отворів. Якщо М. розташовані між кістковими виступами і шкірою, їх скорочення обумовлює зміна шкірного рельєфу.
Всі скелетні, або соматичні (від греч.(грецький) soma — тіло), М. за топографо-анатомічнім принципом можуть бути розділені на М. голови, серед яких розрізняють мімічні і жувальні М., що впливають на нижню щелепу, М. шиї, тулуба і кінцівок. М. тулуба покривають грудну клітку, складають стінки черевної порожнини, унаслідок чого їх ділять на М. грудей, живота і спини. Розчленована скелета кінцівок служить підставою для виділення відповідних груп М.: для верхньої кінцівки — це М. плечового поясу, плеча, передпліччя і кисті; для нижньої кінцівки — М. тазового поясу, стегна, гомілки, стопи.
У людини близько 500 М., пов'язаних із скелетом. Серед них одні великі (наприклад, чотириглава М. стегна), інші — дрібні (наприклад, короткі м'язи спини). Спільна робота М. виконується за принципом синергізму, хоча окремі функціональні групи М. при виконанні певних рухів працюють як антагоністи. Так, спереду на плечі знаходиться двоголова і плечова М., що виконують згинання передпліччя в ліктьовому суглобі, а ззаду розташовується триголова М. плеча, скорочення якої викликає протилежний рух — розгинання передпліччя.
В суглобах кулевидної форми відбуваються прості і складні рухи. Наприклад, в тазостегновому суглобі згинання стегна викликає попереково-клубова М., розгинання — велика сіднична. Стегно відводиться при скороченні середньою і малою сідничних М., а приводиться за допомогою п'яти М. медіальної групи стегна. По кола тазостегнового суглоба локалізуються також М., які обумовлюють обертання стегна всередину і кнаружі.
Найбільш потужні М. розміщуються на тулубі. Це М. спини — випрямляч тулуба, М. живота, складові у людини особливу формацію — черевний прес . У зв'язку з вертикальним положенням тіла М. нижньої кінцівки людини стали сильнішими, оскільки, окрім участі в локомоциі, вони забезпечують опору тіла. М. верхньої кінцівки в процесі еволюції, навпаки, зробилися спритнішими, гарантуючими виконання швидких і точних рухів.
На основі аналізу просторового положення і функціональної діяльності М. сучасна наука користується також наступним їх об'єднанням: група М., що здійснює рухи тулуба, голови і шиї; група М., що здійснює рухи плечового поясу і вільною верхньою кінцівки; М. нижньої кінцівки. В межах цих груп виділяються дрібніші ансамблі.
Ст Ст Купріянов.
Патологія м'язів. Порушення скоротливої функції М. і їх здібності до розвитку і підтримки тонусу спостерігаються при гіпертонії, інфаркті міокарду, міодистрофії, атонії матки, кишечника, сечового міхура, при різних формах паралічів (наприклад, після перенесеного поліомієліту) і ін. Патологічні зміни функцій м'язових органів можуть виникати у зв'язку з порушеннями нервової або гуморальної регуляції, пошкодженнями окремих М. або їх ділянок (наприклад, при інфаркті міокарду) і, нарешті, на клітинному і субклітинному рівнях. При цьому може мати місце порушення обміну речовин (перш за все ферментної системи регенерації макроергічних з'єднань — головним чином АТФ) або зміна білкового скоротливого субстрата. Вказані зміни можуть бути обумовлені недостатнім утворенням м'язових білків на грунті порушення синтезу відповідних інформаційних, або матричних, РНК(рибонуклеїнова кислота), тобто природжених дефектів в структурі ДНК(дезоксирибонуклеїнова кислота) хромосомного апарату кліток. Остання група захворювань, т. о., належить до спадкових захворювань .
Саркоплазматичні білки скелетних і гладких М. представляють інтерес не лише з точки зору можливої участі їх в розвитку в'язкої післядії. Багато хто з них володіє ферментативною активністю і бере участь в клітинному метаболізмі. При пошкодженні м'язових органів, наприклад при інфаркті міокарду або порушенні проникності поверхневих мембран м'язових волокон, ферменти (креатінкиназа лактатдегидрогеназа, альдолаза, амінотрансферази і ін.) можуть виходити в кров. Т. о., визначення активності цих ферментів в плазмі крові при ряду захворювань (інфаркт міокарду, міопатії і ін.) представляє серйозний клінічний інтерес.
Літ.: Енгельгардт Ст А., Ферментативні і механічні властивості білків м'язів, «Успіхи сучасної біології», 1941, т. 14, ст 2; Сент-Джіордьі А., Про м'язову діяльність, пер.(переведення) з англ.(англійський), М., 1947; Іванов І. І., Юрьев Ст А., Біохімія і патобіохимія м'язів, Л., 1961; Поглазов Би. Ф., Структура і функції скоротливих білків, М., 1965; Хайаши Т., Як клітки рухаються, в кн.: Жива клітина, пер.(переведення) з англ.(англійський), 2 видавництва, М., 1966; Хакслі Р., Механізм м'язового скорочення, в сб.(збірка): Молекули і клітки, пер.(переведення) з англ.(англійський), ст 2, М., 1967; Сміт Д., Літальні м'язи комах, там же; Бендолл Дж., М'язи, молекули і рух, пер.(переведення) з англ.(англійський), М., 1970; Арронет Н. І., М'язові і клітинні скоротливі (рухові) моделі, Л., 1971; Льові А., Сикевіц Ф., Структура і функції клітки, пер.(переведення) з англ.(англійський), М., 1971; Іванов І. І., Деякі актуальні проблеми еволюційної біохімії м'язів, «Журнал еволюційної біохімії і фізіології» 1972, т. 8 № 3; Gibbons I. R., The biochemistry of motility, «Annual Review of Biochemistry», 1968, v. 37, р. 521.