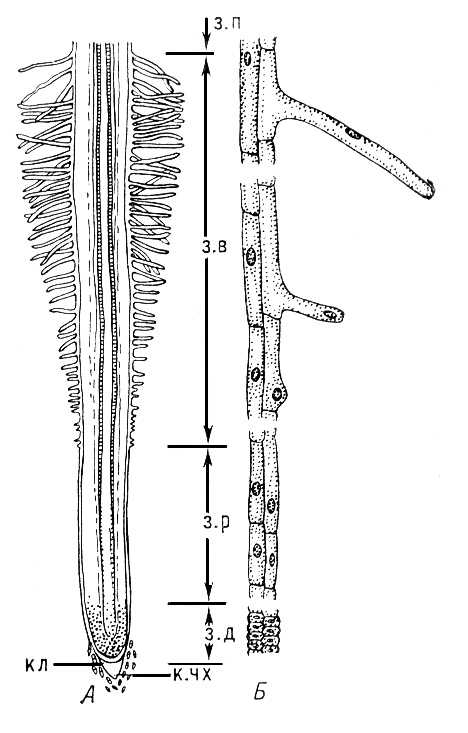
Корінь (radix), один з основних вегетативних органів лістостебельних рослин (за винятком мохів), службовець для прикріплення до субстрата, поглинання з нього води і живильних речовин, первинного перетворення ряду речовин, що поглинаються, синтезу органічних сполук, подальшого переміщення їх в ін. органи рослини, а також для виділення деяких продуктів обміну. В деяких рослин До. несуть додаткові функції, наприклад вмістища запасних речовин, а в корнеотприськових рослин — органу вегетативного розмноження. До. — осьовий орган рослини, за походженням родинний стеблу: у предків наземних рослин при виході їх на сушу на нижньому кінці спорофіту утворилися корневіщеподобниє гілочки — прототип К. Морфологичеськи До. відрізняється від стебла відсутністю листя наявністю чехліка і ендогенним галуженням (виникненням бічних До. у внутрішніх частинах До. — в перициклі ) . Типовий До. має узкоциліндрічеськую або ниткоподібну форму. Зачаток До. існує вже у зародку сім'ю, при проростанні якого він розвивається в головний К. Для До. характерне акропетальне галуження (молодші бічні До. і їх зачатки з'являються ближче до верхівки До.). В багатьох рослин, окрім головного і бічних До., є додаткові. Зовні вони не відрізняються від бічних і несуть ті ж функції, але зазвичай розвиваються на ін. органах рослини — стеблах, листі, підземних і надземних видозмінених втечах (цибулинах, бульбах, кореневищах і т. д.), а також на старих К. Образованіє додаткових До. визначає можливість вегетативного розмноження рослин. Кінчик До. ( мал. 1 ) прикритий кореневим чехліком, що захищає його від пошкоджень. До. зростає в довжину за рахунок ділення клітин верхівковою (апікальной) мерістеми, В т. н. зоні ділення, і значного зростання їх в довжину в зоні зростання, або розтягування, довжина якої зазвичай дорівнює 1—2 мм (інколи до 10 мм ) . За зоною зростання розташована зона поглинання і диференціації; самий поверхневий шар кліток цієї зони диференціюється в епіблему, клітки якої утворюють кореневі волоски, поглинання, що збільшують поверхню, До. у 5—20 разів. Диференціація мерістеми в первинні постійні тканини починається в зоні зростання, але виразно виявляється в зоні поглинання і в т.з. зоні проведення. Первинна анатомічна будова До. всіх насінних рослин схоже ( мал. 2 ). Під шаром кліток епіблеми розташована багатошарова первинна кора, яка складається з тонкостінних живих паренхімних клітин. В деяких рослин оболонки 1—3 зовнішніх шарів кліток кори (екзодерма ) опробковевают і після відмирання епіблеми беруть на себе захисну функцію. Самий внутрішній шар первинної кори (ендодерма ) складається з одного шару кліток, оболонки яких частково опробковевают або одревесневают і незрідка товщають; лише деякі клітки (т.з. пропускні) залишаються мало зміненими. Ендодерма оточує центральний циліндр, або стель, К. Одін або декілька зовнішніх шарів його кліток утворюють перицикл, тонкостінні живі клітини якого довго зберігають мерістематічеськую активність; у нім закладаються бічні До., додаткові нирки; при його участі здійснюється вторинне потовщення До. Велику частину центрального циліндра займає складний провідний радіальний пучок, в якому чергуються елементи ксилема і флоеми . В однодольних рослин первинна анатомічна будова До. зберігається все життя, в дводольних і голосеменних воно змінялося вторинним ( мал. 3 ). При цьому в центральному циліндрі пучковий камбій відкладає (між флоемою і ксилемою) до центру вторинну ксилему (деревину), а до периферії — вторинну флоему (луб). З межпучкового камбію, що знаходиться проти променів первинної ксилеми в перициклі, утворюється паренхіма первинного серцевинного (лубо-деревінного) променя, а пробковий камбій (фелоген) відкладає кнаружі клітки пробки; вона стає покривною тканиною повторно потовщеного До., а вся первинна кора скидається. Ці зміни обумовлюють розростання До. у товщину. Розрізняють т.з. ростові До., які швидко зростають (у сприятливих умовах — в середньому від 1 до 3 см в добу), товщають, рано опробковевают, складають скелет кореневої системи і у ряді випадків забезпечують вегетативне розмноження рослини, і що «смокчуть» До. — тонкі, ніжні, короткі, повільно зростаючі, переважно недовговічні.
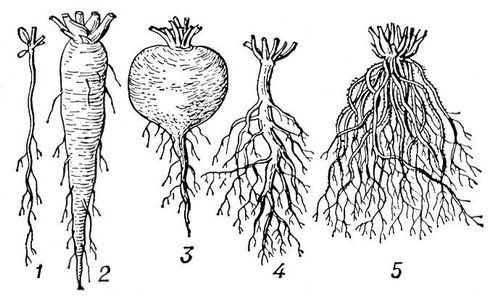
Сукупність До. однієї рослини називається кореневий системою, загальну форму і характер якої визначає співвідношення зростання головного, бічних і додаткових До. ( мал. 4 ). При переважаючому зростанні головного До. утворюється стрижньова коренева система (люпин, бавовник); при слабкому зростанні або ранньому відмиранні головного і переважаючому розвитку великого числа додаткових До. утворюється мочкувата коренева система (однодольні, з дводольних — види жовтця, подорожника і ін.). До. в хлібних злаків (іржі, пшениці) проникають на глибину 1—1,5 м-коду, в люцерни —до 10 м-кодів, в деревних рослин можуть заглиблюватися на 10—12 м-коду, хоча зазвичай твердий грунт перешкоджає проникненню їх глибше за 3—5 м-код; площа, зайнята кореневою системою однієї рослини хлібних злаків, досягає в поперечнику 40—60 см, гарбузи — 6—8 м-код, в деревних рослин зазвичай у декілька разів перевищує діаметр крони, досягаючи 10—18 м. Сумарна довжина До. однорічного рослини може складати декілька кілометрів, а загальна поверхня До., враховуючи поверхню волосків, у багато разів перевищує поверхню надземних органів. По розташуванню в грунті залежно від місця існування розрізняють кореневі системи спеціалізовані (поверхневі або глибинні) і універсальні, такі, що рівномірно розвиваються вшир і углиб. Наприклад, в північній лісовій зоні на підзолистих грунтах, що часто пересичених вологою, погано аеруються, з важко проникною підгрунтям, коренева система рослин на 90—95% зосереджена в поверхневих шарах (10—15 см ) . В зоні напівпустель і пустель в одних рослин є лише поверхневі До., які використовують осідання ранньої весни (ефемери ) або вологу дощів і конденсаційну вологу, що осідає у верхніх шарах грунту в нічний час (кактуси); в інших До. досягають грунтових вод (у верблюжої колючки). Універсальні кореневі системи використовують в різний час вологу різних горизонтів, унаслідок чого рослини можуть вегетіровать все літо (джузгун, саксаул, ефедра). У зонах недостатнього зволоження яскраво виявляється ярусність в розташуванні До. різних видів рослин, що виростають спільно на одній площі. Співвідношення між До. і надземними частинами рослин також залежить від грунтово-кліматичних умов. У північній вологій зоні маса кореневої системи в 5—10 разів поступається масі надземних частин. Через До. рослини поглинають з грунту головним чином іони мінеральних солей, а також деякі продукти життєдіяльності грунтових мікроорганізмів і кореневі виділення ін. рослин. Поглинені До. з'єднання азоту, фосфору і сірки взаємодіють з продуктами фотосинтезу, що притікають з листя, з утворенням амінокислот, нуклеотидів і ін. органічних сполук. По судинах ксилеми елементи у формі іонів (калій, кальцій, магній, фосфор) або органічних молекул (азот, сірка) в результаті дії кореневого тиску і транспірування пересуваються в листя і стебла. У ДО. синтезуються також алкалоїди (наприклад, нікотин), гормони зростання (киніни, гибберелліни ) і ін. фізіологічно активні речовини. В деяких рослин, переважне сімейство бобів, в До. поселяються бактерії, що викликають утворення бульб (з паренхіми До.). До. рослин виділяють в грунт іони мінеральних солей, амінокислоти, цукру, сечовину, фенольні з'єднання і ін. В одних рослин (наприклад, сімейство пасльонових) До. виділяють також ауксини і речовини типа гиббереллінов, стимулюючі зростання рослин, в ін. — студнеобразниє речовини, створюючі корнезащитниє чохли. Виділення До. сприяють розвитку в різосфері грунтових мікроорганізмів. З віком рослини кількість виділень і чисельність різосферних мікроорганізмів зменшуються. Клубеньковиє бактерії грають важливу роль в азотному живленні рослин і підвищенні родючості грунту. На поверхні або усередині До. багатьох деревних і трав'янистих рослин поселяються гриби, утворюючи т.з. мікоризу ; в таких випадках кореневі волоски на До. не розвиваються.
На До. багатьох рослин утворюються додаткові нирки, що дають надземні втечі (в корнеотприськових рослин), В ряду рослин До. служать местомом відкладення запасних живильних речовин (див. Коренеплоди ). В деяких дерев тропічних лісів поблизу підстави стволів або від гілок відходять бічні або додаткові До. — доськовідниє ходульні, столбовідниє, службовці для опори і живлення. В тих, що лазять ліан (наприклад, в плюща) розвиваються коріння-причіплювання. У тропіках багато епіфіти утворюють додаткове повітря До., що поглинають через багатошарову поверхневу тканину (т.з. веламен) воду з атмосферних опадів. В деяких тропічних епіфітних орхідей, а також в рослин сімейства подостемонових листя і стебла недорозвинені і їх вегетативні органи представлені головним чином плоскими зеленими До., що виконують функції асиміляції. В рослин, що живуть на бідних киснем грунтах (наприклад, в болотяного кипариса, в мангрових дерев і ін.), є дихательниє До., або пневматофори, верхівки яких розташовані над грунтом або водою і забезпечують підземні органи повітрям. В деяких пальм і рослин сімейства маренових частина горизонтальних До. перетворюється на захисних колючки. До. паразитуючих на деревах рослин, наприклад омели, мають вигляд довгих циліндрових тяжей, розташованих в корі дерева. В растеній—паразітов (вовчок, повіліка і ін.) і напівпаразитів (марьянник, дзвінець і ін.) коренева система розвивається слабо; при цьому закінчення деяких До. упроваджуються у вигляді особливих присосків ( гаусторій ) в тіло рослини-господаря, висмоктуючи з нього живильні речовини. В деяких рослин (наприклад, в роголістника, пухирчатки і ін.) До. відсутні, що пов'язане із специфічними умовами їх існування. Про функцію До. див.(дивися) також Водний режим рослин, Мінеральне живлення рослин .
До. багатьох рослин широко використовуються людиною. Вони мають велике харчове і господарське значення. До., що містять крохмаль, цукру масла, алкалоїди, гутаперчу, фарбувальні і ін. коштовні речовини, застосовуються в медицині і промисловості. Рослини з могутньо розвиненою кореневою системою використовуються для закріплення рухливих пісків, ярів і еродованих грунтів.
Літ.: Красовськая І. Ст, Огляд робіт по морфології і фізіології коріння, «Тр. по прикладній ботаніці генетиці і селекції», 1928, т. 18, ст 5; її ж, Закономірності будови кореневої системи хлібних злаків «Ботанічний журнал», 1950, т. 35 № 4; Шалит М. С., Підземна частина деяких лугових, степових і пустинних рослин і фітоценозів, «Праці Ботанічного інституту АН(Академія наук) СРСР. Сірок. 3, Геоботаніка», 1950, ст 6; Сабінін Д. А., Фізіологічні основи живлення рослин, М., 1955; Качинський Н. А., Грунт, її властивості і життя, М., 1956; Колісників Ст А., Коренева система плодових і ягідних рослин і методи се вивчення, М., 1962; Федоров А. А., Цегельників М. Е. і Артюшенко З. Т., Атлас по описовій морфології вищих рослин, [т. 2], М.— Л., 1962; Колосов І. І., Поглинювальна діяльність кореневих систем рослин, М., 1962; Рахтєєнко І. Н., Зростання і взаємодія кореневих систем деревних рослин, Мінськ, 1963; Воронін Н. С., Еволюція первинних структур в корінні рослин, «Уч. зап.(західний) Калузького педагогічного інституту», 1964, ст 13; Жваво Л. А., фізіологія кореневої системи рослин в умовах засолення, Л., 1969; Есау До., Анатомія рослин, пер, з англ.(англійський), М., 1969.
О. Н. Чистякова, Р. П. Барикина, Д. Би. Вахмістрів.