Земноводні, амфібії (Amphibia) (від греч.(грецький) amphíbios — що живе двояким життям), клас хребетних тварин. З. — перші хребетні, такі, що перейшли від водного до водно-наземного способу життя. Відкладають ікру, подібно до риб, т.к. іх яйця (ікра) і зародки позбавлені пристосувань для наземного розвитку (анамнії ) . Розвиток завершується метаморфозом, під час якого личинки втрачають схожість з рибами і перетворюються на дорослих тварин. Організація З. як наземних хребетних багато в чому недосконала: інтенсивність обміну речовин дуже невисока, температура тіла непостійна і відповідає температурі зовнішнього середовища.
До сучасних З. відноситься близько 2850 видів, що об'єднуються в 3 загони: безногі, хвостаті і безхвості З. Безногі земноводні мають подовжене тіло, кінцівки і хвіст відсутні. Хвостаті земноводні мають витягнуте тіло, хвіст добре розвинений, ноги зазвичай слабкі і короткі. Безхвості земноводні мають дві пари кінцівок, по землі пересуваються стрибками; у воді плавають, відштовхуючись задніми кінцівками, забезпеченими у більшості видів плавальні перетинками.
М'яка і волога шкіра грає у З. важливу роль в диханні. Необхідна для газообміну вологість шкіри підтримується виділеннями слизистих залоз. На спинній стороні тіла розташовані крупні білкові (серозні) залози, виділення яких отруйні. Лише в деяких безногих З. у шкірі є дрібні кісткові лусочки. Потиличних виростків два. Верхній кістковий елемент під'язикової дуги перетворений на слухову кісточку. Під час метаморфоза зяброві дуги редукуються і перетворяться разом з нижніми елементами під'язикової дуги в під'язикову кість. Загальне число хребців вагається від 9 (у більшості безхвостих З.) до 200 і більш (в безногих З.). У більшості безхвостих З. ребра повністю зредуковані. Кінцівки парні, п'ятипалого типа. Клубові кістки тазу (сильно подовжені в безхвостих З.) зчленовуються з поперечними відростками крижового хребця.
Головний мозок З. має добре розвинений передній мозок, півкулі якого повністю розділені. Мозочок розвинений дуже слабо. Черепномозкових нервів 10 пар. В личинок З. є органи бічної лінії (див. Бічні органи ). Акомодація ока здійснюється переміщенням кришталика. В тих, що живуть в грунті безногих З. і в печерних хвостатих З. очі недорозвинені. Безхвості З., окрім внутрішнього вуха, мають порожнину середнього вуха і барабанну перетинку. Органи дотику добре розвинені. Органи нюху добре виражені в безногих і хвостатих З. Органи смаку розвинені слабо.
В дорослому стані все З. харчуються виключно тваринною їжею. Зуби служать лише для схоплювання і утримання видобутку. В жаб і піп зуби повністю відсутні. На відміну від риб, З. мають рухливу мову, багату залозами, що виділяють клейкий слиз, що сприяє захопленню дрібного видобутку. Травний тракт З. порівняно короткий. Більшість З. у дорослому стані має легені, хоани (внутрішні ніздрі) і горлові хрящі. У зв'язку з відсутністю грудної клітки в більшості З. при опусканні дна ротової порожнини повітря всмоктується в неї через ніздрі; потім ніздрі замикаються клапанами, а дно рота піднімається до піднебіння, нагнітаючи повітря в легені через горлову щілину. Легеневе дихання доповнюється шкіряним, яке інколи має переважаюче значення (наприклад, в безлегеневих саламандр ). Хвостаті З., що мешкають у воді, дихають зябрами, зберігаючи їх протягом всього життя.
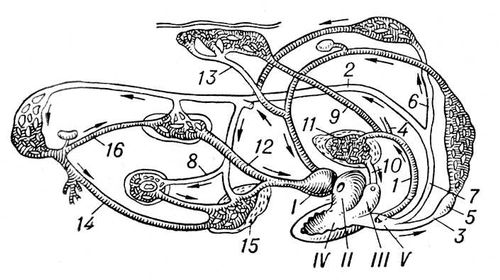
Кровообіг личинок З. схоже з кровообігом риб. Серце З. після метаморфоза набуває трикамерної будови, тобто утворено 2 передсердям і 1 шлуночком. Праве передсердя отримує венозну кров, ліве — артеріальну (з легенів і шкіри). Венозна і артеріальна кров лише частково змішуються в порожнині шлуночку, стінки якого мають складну систему мускульної щаблини. У легеневі вени прямує головним чином венозна кров, дуги аорти заповнюються змішаною кров'ю, і лише сонні артерії отримують артеріальну кров. (мал. 1.)
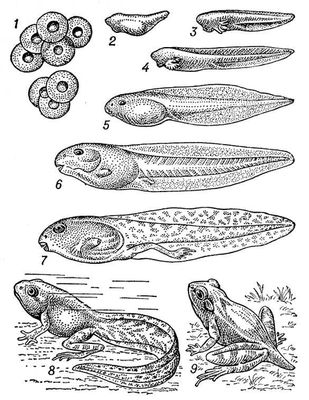
У дорослих З. є парна тулубова брунька. В личинок на ранніх стадіях розвитку функціонує т.з. головна брунька, або передбрунька. Велике значення у водному обміні має сечовий міхур. У самців є парні семенникі, у самок — парні яєчники. Яйцеклітини виводяться назовні через яйцепроводи, що впадають в клоаку . Статевозрілості З. досягають найчастіше до 3—4-го року. Більшість З. відкладає ікру у водоймища. В період розмноження для безхвостих З. характерні «весняні концерти» (спів самців), для хвостатих З. — шлюбні ігри; в багатьох форм помітно посилюється статевий диморфізм. Майже у всіх безхвостих і небагато хвостатих З. запліднення зовнішнє, у більшості хвостатих і в безногих З. —внутреннєє. Лише небагато З. живородящі. Всі інші відкладають ікру. Плодючість З. вагається від 3 ікринок до 28 тис. Личинка в безхвостих З. (називається пуголовком ) особливо сильно відрізняється від дорослої особини. При метаморфозі зовнішні зябра зникають, органи чуття перебудовуються відповідно умовам повітряного середовища; у епідермісі розвивається роговий шар. В З., що відкладають ікру на суші, інколи спостерігається перехід до розвитку без метаморфоза, тобто прямий розвиток. Личинки деяких хвостатих З. можуть досягати статевозрілості до метаморфоза (неотенія ) . Більшість З., відклавши ікру, покидає її. В деяких форм батьки переносять на собі памолодь або ікру (наприклад, самці жаби-повитухи). Деякі З. виношують памолодь, наприклад самець рінодерми Дарвіна, самки сумчастих жаб і піпи. (Мал. 2 і 3.)
Більшість З. корисні, т.к. унічтожают шкідників сільського господарства і служать їжею для ін. тварин. У багатьох країнах (Франція, Італія, США і ін.) деякі жаби уживаються людиною в їжу. Деякі З., наприклад жаби, — класичні об'єкти для фізіологічних експериментів.
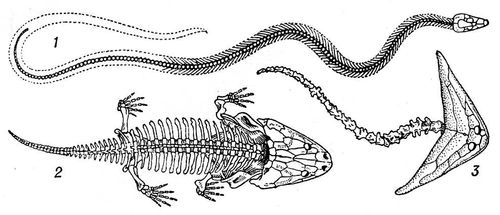
Копалини З. набагато багаточисельно і всілякіше сучасних. Класифікація копалин З. заснована на будові їх хребта і черепа. Найбільш древні і примітивні З. — іхтіостеги — володіли значною схожістю з кистеперимі рибами, будучи їх нащадками. Головна гілка копалин З. складали лабірінтодонти ; від них, мабуть, відокремилися батрахозаври, безхвості і лепоспондільниє З., а також мікрозаври. Батрахозаври — вірогідні предки плазунів . З'явившись в девоне, З. досягли широкого поширення і різноманітності в карбоні, чому сприяв вологий і жаркий клімат того періоду; у пермський, сухіший, період серед З. з'явилися багато наземних форм. У тріасі різноманіття З., представлених переважно водними формами, різко скоротилося; 2-й етап їх відносного процвітання доводиться вже на кайнозой. Деякі копалини З. досягали гігантських розмірів, маючи череп завдовжки більше 1 м-код (мастодонзавр ) . Основні місцезнаходження залишків древніх З. (девон — тріас) відомі з Північної півкулі, окремі знахідки — з Південної Африки і Індії. У СРСР залишки копалин З. багаточисельні в східній частині Російської платформи і служать надійними керівними формами для встановлення геологічного віку відкладень. (Мал. 4 і 5.)
Літ.: Ефремов І. А., Вьюшков Би. П., Каталог місцезнаходження пермських і тріасах наземних хребетних на території СРСР, «Тр. Палеонтологічного інституту АН(Академія наук) СРСР», 1955, т. 46; Терентьев П. Ст, Герпетологія М., 1961; Шмальгаузен І. І., Походження наземних хребетних, М., 1964; Основи палеонтології. Земноводні, плазуни і птиці, М., 1964; Життя тварин, т. 4, ч. 2, М., 1969; Noble G., The biology of the amphibia, N. Y. — L., 1931; Angel F., Vie et moeurs des amphibiens. P., 1947; Romer A. S., Review of Labyrinthodontia, «Bulletin Museum Comparative Zoology», 1947, v. 99 № 1; Physiology of the amphibia, ed. J. A. Moore, N. Y., 1964.