Осмотичний тиск , дифузний тиск, термодинамічний параметр, що характеризує прагнення розчину до пониження концентрації при зіткненні з чистим розчинником унаслідок зустрічної дифузії молекул розчиненої речовини і розчинника. Якщо розчин відокремлений від чистого розчинника напівпроникною мембраною, то можлива лише однобічна дифузія — осмотичне всмоктування розчинника через мембрану в розчин. В цьому випадку О. д. стає доступною для прямого виміру величиною, рівною надлишковому тиску, прикладеному з боку розчину при осмотичній рівновазі (див. Осмос ). О. д. обумовлено пониженням хімічного потенціалу розчинника у присутності розчиненої речовини. Тенденція системи вирівнювати хімічні потенціали у всіх частинах свого об'єму і перейти в стан з нижчим рівнем вільній енергії викликає осмотичне (дифузійний) перенесення речовини. О. д. у ідеальних і гранично розбавлених розчинах не залежить від природи розчинника і розчинених речовин; при постійній температурі воно визначається лише числом «кінетичних елементів» — іонів, молекул, ассоциатов або колоїдних часток — в одиниці об'єму розчину. Перші виміри О. д. виробив В.Пфеффер (1877), досліджуючи водні розчини тростинного цукру. Його дані дозволили Я. X.Вант-Гоффу встановити (1887) залежність О. д. від концентрації розчиненої речовини, співпадаючу формою з Бойля — Маріотта законом для ідеальних газів. Виявилось, що О. д. (p) чисельно дорівнює тиску, який надав би розчинену речовину, якби воно при даній температурі знаходилося в стані ідеального газу і займало об'єм, рівний об'єму розчину. Для вельми розбавлених розчинів недиссоціюючих речовин знайдена закономірність з достатньою точністю описується рівнянням: pv = nrt , де n — число мілі розчиненої речовини в об'ємі розчину V; R — універсальна газова постійна; Т — абсолютна температура. В разі дисоціації речовини в розчині на іони в праву частину рівняння вводиться множник i > 1, коефіцієнт Вант-Гоффа; при асоціації розчиненої речовини i < 1. О. д. реального розчину (p'') завжди вище, ніж ідеального (p''''), причому відношення p''/ p'''' = g , зване осмотичним коефіцієнтом, збільшується із зростанням концентрації. Розчини з однаковим О. д. називається ізотонічними або ізоосмотичними. Так, різні кровозамінники і фізіологічні розчини ізотонічни відносно внутрішніх рідин організму. Якщо один розчин порівняно з іншим має вище О. д., його називають гіпертонічним, а що має нижче О. д. — гіпотонічним.
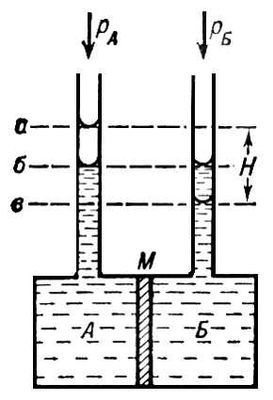
О. д. вимірюють за допомогою спеціальних приладів — осмометрів. Розрізняють статичні і динамічні методи виміру. Перший метод заснований на визначенні надлишкового гідростатичного тиску по висоті стовпа рідини Н в трубці осмометрів ( мал. ) після встановлення осмотичної рівноваги при рівності зовнішнього тиску p A і р Би в камерах А і Б. Второй метод зводиться до виміру швидкостей v всмоктування і витискування розчинника з осмотичного вічка при різних значеннях надлишкового тиску D p = p A — р Би з подальшою інтерполяцією отриманих даних до n = 0 при D p = р. Багато осмометрів дозволяють використовувати обидва методи. Одна з головних труднощів у вимірі О. д. — правильний підбір напівпроникних мембран. Зазвичай застосовують плівки з целофану, природних і синтетичних полімерів, пористі керамічні і скляні перегородки. Вчення про методи і техніку виміру О. д. називаються осмометрією. Основний додаток осмометрії — визначення молекулярної маси ( М-код ) полімерів. Значення М-код обчислюють із співвідношення, де з — концентрація полімеру по масі; А — коефіцієнт, залежний від будови макромолекули.
О. д. може досягати значних величин. Наприклад, 4%-ний розчин цукру при кімнатній температурі має О. д. близько 0,3 Мн/м-код 2 , а 53%-ний — близько 10 Мн/м-код 2 ; О. д. морської води близько 0,27 Мн/м-код 2 .
Л. А. Шиц.
О. д. у клітках тварин, рослин, мікроорганізмів і в біологічних рідинах залежить від концентрації речовин, розчинених в їх рідких середовищах. Сольовий склад біологічних рідин і кліток, характерний для організмів кожного виду, підтримується виборчою проникністю біологічних мембран для різних солей і активним транспортом іонів. Відносна постійність О. д. забезпечується водно-сольовим обміном, тобто всмоктуванням, розподілом, вжитком і виділенням води і солей (див. Виділення, Видільна система, Осморегуляція ). В т.з. гіперосмотичних організмів внутрішнього О. д. більше зовнішнього, в гипоосмотічеських — менше зовнішнього; в ізоосмотичних (пойкилоосмотічеських) внутрішнє О. д. дорівнює зовнішньому. У першому випадку нони активно поглинаються організмом і затримуються в нім, а вода поступає через біологич. мембрани пасивно, відповідно до осмотичного градієнта. Гіперосмотична регуляція властива прісноводим організмам, мор.(морський) хрящовим рибам (акули, скати) і всім рослинам. В організмів з гипоосмотічеськой регуляцією є пристосування для активного виділення солей. У костистих риб переважаючі в океанічних водах іони Na + і Cl — виділяються через зябра, у морських плазунів (змії і черепахи) і у птиць — через особливі сольові залози, розташовані в області голови. Іони Mg 2 + ,, в цих організмів виділяються через нирки. О. д. в гипер- і гипоосмотічеських організмів може створюватися як за рахунок іонів, переважаючих в зовнішньому середовищі, так і продуктів обміну. Наприклад, у акулових риб і скатів О. д. на 60% створюється за рахунок сечовини і тріметіламмонія; у плазмі крові ссавців — головним чином за рахунок іонів Na + і Cl — ; у личинках комах — за рахунок всіляких низькомолекулярних метаболітов. В морських одноклітинних, голкошкірих, головоногих молюсків, міксин і ін. ізоосмотичних організмів, в яких О. д. визначається О. д. зовнішнього середовища і дорівнює йому, механізми осморегуляції відсутні (виключаючи клітинні).
Діапазон середніх величин О. д. у клітках організмів, не здатних підтримувати осмотичний гомеостаз, досить широкий і залежить від вигляду і віку організму, типа кліток і О. д. довкілля. У оптимальних умовах О. д. клітинного соку наземних органів болотяних рослин вагається від 2 до 16 ат , в степових — від 8 до 40 ат . У різних клітках рослини О. д. може різко розрізнятися (так, в мангрових О. д. клітинного соку близько 60 ат , а О. д. у судинах ксилеми не перевищує 1—2 ат ). В гомойосмотічеських організмів, тобто здатних підтримувати відносну постійність О. д., середньої величини і діапазон коливань О. д. різні (дощовий черв'як — 3,6—4,8 ат , прісноводі риби — 6,0—6,6, океанічні костисті риби — 7,8—8,5, акуловиє — 22,3—23,2, ссавці — 6,6—8,0 ат ). У ссавців О. д. більшості біологічних рідин рівне О. д. крові (виняток становлять рідини, що виділяються деякими залозами, — слина, піт, мочивши і ін.). О. д., створюване в клітках тварин високомолекулярними з'єднаннями (білки, полісахариди і ін.), трохи, але грає важливу роль в обміні речовин (див. Онкотичний тиск ).
Ю. Ст Наточин, Ст Ст Кабанів.
Літ.: Мелвін-Хьюз Е. А., Фізична хімія, пер.(переведення) з англ.(англійський), кн. 1—2, М., 1962; Курс фізичної хімії, під ред. Я. І. Герасимова, т. 1—2, М. — Л., 1963—1966; Пасинський А. Р., Колоїдна хімія, 3 видавництва, М., 1968: Проссер Л., Браун Ф., Порівняльна фізіологія тварин, пер.(переведення) з англ.(англійський), М., 1967; Гриффін Д., Новік Ел., Живий організм, пер.(переведення) з англ.(англійський), 1973; Нобел П., Фізіологія рослинної клітки (физико-хімічний підхід), пер.(переведення) з англ.(англійський), М., 1973.